You might also like
- Flujo de Información GenéticaDocument23 pagesFlujo de Información GenéticaKennyDeLosReyesNo ratings yet
- Abbu Laba Di BaDocument5 pagesAbbu Laba Di Bammamerto2005No ratings yet
- Assignment For The Course Molecular Biology (Cheg 6181) On Regulation of Gene ExpressionDocument40 pagesAssignment For The Course Molecular Biology (Cheg 6181) On Regulation of Gene ExpressionBro Gi GisheNo ratings yet
- Ans 201 Anatomy and Physiology of Farm AnimalsDocument33 pagesAns 201 Anatomy and Physiology of Farm AnimalsAdewaleNo ratings yet
- Endoplasmic ReticulumDocument5 pagesEndoplasmic ReticulumAbdul RehmanNo ratings yet
- 1 - Give An Illustrated Account of The Fine Staracture of A Plant Cell and Describe The Function of The Diffrent Cell OrgnagesDocument19 pages1 - Give An Illustrated Account of The Fine Staracture of A Plant Cell and Describe The Function of The Diffrent Cell OrgnagesMD shah KhanNo ratings yet
- Pys PDFDocument5 pagesPys PDFMD shah KhanNo ratings yet
- Genome Control and Cell VariationDocument25 pagesGenome Control and Cell VariationwbaturleyNo ratings yet
- If You Understand How Proteins WorkDocument4 pagesIf You Understand How Proteins WorkMah Song WeiNo ratings yet
- Lesson 2: Molecular Organization of CellsDocument12 pagesLesson 2: Molecular Organization of CellsremshopNo ratings yet
- Cell Organelles and Their FunctionsDocument8 pagesCell Organelles and Their FunctionsPrincess Angie GonzalesNo ratings yet
- Unit: 07: Mechanism of Cellular Differentiation / Differential Gene ExpressionDocument76 pagesUnit: 07: Mechanism of Cellular Differentiation / Differential Gene ExpressionMuhammad SibtainNo ratings yet
- 04 Structure To FunctionDocument5 pages04 Structure To Function897291868100% (1)
- Gene Concept EvolutionDocument20 pagesGene Concept Evolutionshahena sNo ratings yet
- Eukaroyotic CellDocument5 pagesEukaroyotic CellMa. Kate Nicole D. PadpadNo ratings yet
- The Basics of Cell Life with Max Axiom, Super Scientist: 4D An Augmented Reading Science ExperienceFrom EverandThe Basics of Cell Life with Max Axiom, Super Scientist: 4D An Augmented Reading Science ExperienceNo ratings yet
- PHYSIOLOGYDocument16 pagesPHYSIOLOGYIqtader hoque khan AkibNo ratings yet
- Smooth and Granular Endoplasmic Reticulum RibosomesDocument26 pagesSmooth and Granular Endoplasmic Reticulum RibosomesArfiNo ratings yet
- 6.1-How Are Cells Studied?: Magnification Resolution Contrast Electron Microscope (EM)Document7 pages6.1-How Are Cells Studied?: Magnification Resolution Contrast Electron Microscope (EM)Sadasivarao GaddeNo ratings yet
- Kanzul Cell & Molecular BiologyDocument12 pagesKanzul Cell & Molecular BiologyMuhammad FarhanNo ratings yet
- Cells - Molecules and MechanismsDocument277 pagesCells - Molecules and MechanismsscottswallowsNo ratings yet
- Gene Regulation - Operon Theory - Microbiology - OpenStaxDocument12 pagesGene Regulation - Operon Theory - Microbiology - OpenStaxAleksandra Sanja MartinovicNo ratings yet
- Cellmol-Group12-Final - PPTX 20231023 073441 0000Document29 pagesCellmol-Group12-Final - PPTX 20231023 073441 0000Noronisa PendalidayNo ratings yet
- bbt193003 - Yeshua ShahzadDocument9 pagesbbt193003 - Yeshua ShahzadAbuzar KhanNo ratings yet
- Plant CellDocument8 pagesPlant CellIrish Nicole PacionNo ratings yet
- Gene Protein and RegulationDocument4 pagesGene Protein and Regulationapi-248290141No ratings yet
- Cell Biology 1Document1 pageCell Biology 1adedapo adedireNo ratings yet
- BIO230 - Section 1 Regulation of Genome Expression Lecture 1-9Document36 pagesBIO230 - Section 1 Regulation of Genome Expression Lecture 1-9yusrawasim147No ratings yet
- 0-Introduction NotesDocument7 pages0-Introduction Notesjimmy.bousaba27No ratings yet
- Cells, Organelles, Microscopy TechniquesDocument34 pagesCells, Organelles, Microscopy TechniquesJeffrey100% (1)
- Molecular BiologyDocument9 pagesMolecular Biologyrifat RasheedNo ratings yet
- As OCR Biology Revision PackDocument28 pagesAs OCR Biology Revision PackJoel TrelivingNo ratings yet
- The Cell: Anabolism CatabolismDocument14 pagesThe Cell: Anabolism CatabolismHashim GhazoNo ratings yet
- Gene Expression - Learn Science at ScitableDocument5 pagesGene Expression - Learn Science at ScitableIshmeet KaurNo ratings yet
- Genetics VanshDocument21 pagesGenetics VanshJatinNo ratings yet
- Gene ExpressionDocument16 pagesGene ExpressionmidhunbonumahanthiNo ratings yet
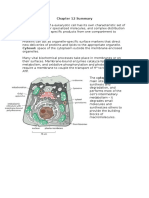
- Chapter 12 SummaryDocument7 pagesChapter 12 SummaryCharlotteNo ratings yet
- Micro .3.Document13 pagesMicro .3.Hashim GhazoNo ratings yet
- Homework 1 &2 Biology IUDocument5 pagesHomework 1 &2 Biology IUVanessa NguyenNo ratings yet
- M4 Check-In Activity 1 - VenusDocument2 pagesM4 Check-In Activity 1 - VenusGrezha Mari VenusNo ratings yet
- Regulation of Gene Expression in Eukaryotes: Md. Murad Khan Lecturer Dept. of Microbiology Jagannath UniversityDocument68 pagesRegulation of Gene Expression in Eukaryotes: Md. Murad Khan Lecturer Dept. of Microbiology Jagannath UniversityCharls John ErcilloNo ratings yet
- NotesDocument37 pagesNotesDaideepya SaxenaNo ratings yet
- Cellbiologyreview AnswersDocument8 pagesCellbiologyreview AnswersCARLISABEL GALLARDONo ratings yet
- Topic 3 Cell Differentiation and VariationDocument7 pagesTopic 3 Cell Differentiation and VariationdaniaNo ratings yet
- How To Approach Bioinformatics Analysis in Protein Structure and FunctionDocument4 pagesHow To Approach Bioinformatics Analysis in Protein Structure and FunctionSuperSonic2422No ratings yet
- Morphological Function of The Cell Presentation PHDocument52 pagesMorphological Function of The Cell Presentation PHDoc HamsNo ratings yet
- Cell Organelles and Their FunctionsDocument24 pagesCell Organelles and Their FunctionsWina MaeNo ratings yet
- 17:10 Lezione 7Document13 pages17:10 Lezione 7gaiaadamo15No ratings yet
- Cell Structures: The BasicsDocument4 pagesCell Structures: The BasicsParama CinthyaNo ratings yet
- Polyribosomes & Posttranslational EventsDocument24 pagesPolyribosomes & Posttranslational Eventsahmed mediaNo ratings yet
- Cell Organelles - Structure and FunctionDocument6 pagesCell Organelles - Structure and FunctionKaryl Marie Rodas JaminNo ratings yet
- CH 6 BiologyDocument4 pagesCH 6 BiologysophieowensNo ratings yet
- Differences between prokaryotic and eukaryotic cellsDocument18 pagesDifferences between prokaryotic and eukaryotic cellssalahuddin_md5935100% (1)
- ZoologyDocument2 pagesZoologygabrf.pascualNo ratings yet
- Circuit Equivalent of A CellDocument3 pagesCircuit Equivalent of A Cellmarosep123No ratings yet
- Basic Cell Structure & FunctionDocument27 pagesBasic Cell Structure & FunctionJawaad AsifNo ratings yet
- Bio 1Document34 pagesBio 1mtanveerhasan2001No ratings yet
- Human Chromosomes: An Illustrated Introduction to Human CytogeneticsFrom EverandHuman Chromosomes: An Illustrated Introduction to Human CytogeneticsRating: 5 out of 5 stars5/5 (1)
- Youcai-Chenglong2017 Book PollutionControlAndResourceReuDocument447 pagesYoucai-Chenglong2017 Book PollutionControlAndResourceReuMartin DuarteNo ratings yet
- 4.2 Procesos de Pulpeo - Pulpeo QuímicoDocument38 pages4.2 Procesos de Pulpeo - Pulpeo QuímicoMiguel MontielNo ratings yet
- Stepan Formulation 1220Document2 pagesStepan Formulation 1220Devesh KumarNo ratings yet
- Unit 1 ElectrochemistryDocument73 pagesUnit 1 ElectrochemistrySwaasNo ratings yet
- HF - H2oDocument4 pagesHF - H2ovasudev palaganiNo ratings yet
- Exolit AP 422Document3 pagesExolit AP 422محمد عزتNo ratings yet
- List of Definition in Chemistry F4Document7 pagesList of Definition in Chemistry F4Lil' Nyeh100% (1)
- Vietnam Regulations on Environmental StandardsDocument14 pagesVietnam Regulations on Environmental Standardsrequirement214No ratings yet
- XI Chemistry Chapter Equilibrium Key ConceptsDocument10 pagesXI Chemistry Chapter Equilibrium Key ConceptsSridhar MarellaNo ratings yet
- Al Harrasi Dan Al Saidi 2008 Phytochemical Analysis of The Essential Oil From Botanically Certified Oleogum Resin of BoswelliaDocument9 pagesAl Harrasi Dan Al Saidi 2008 Phytochemical Analysis of The Essential Oil From Botanically Certified Oleogum Resin of BoswelliaYuliana Sabarina LewarNo ratings yet
- Mass Spectrometry (Multiple Choice) QPDocument12 pagesMass Spectrometry (Multiple Choice) QPYusuf AlamNo ratings yet
- Science 9 - Q2 - Mod3 - IONS-HOW-ARE-THEY-FORMED - VerFinalDocument23 pagesScience 9 - Q2 - Mod3 - IONS-HOW-ARE-THEY-FORMED - VerFinalAbel Emmanuel Solitario CabralesNo ratings yet
- SEM Petrology AtlasDocument247 pagesSEM Petrology AtlasRicardo Percilio100% (1)
- Extraction of Oil From Oil Cake by Soxhlet ExtractorDocument11 pagesExtraction of Oil From Oil Cake by Soxhlet Extractormahbub1332No ratings yet
- Indicators and PH ScaleDocument3 pagesIndicators and PH ScaleToni - Ann IrvingNo ratings yet
- Aerospace Material Specification: Plating, Nickel General PurposeDocument8 pagesAerospace Material Specification: Plating, Nickel General PurposeSURYAS63No ratings yet
- Analytical Chemistry Laboratory Manual.Document43 pagesAnalytical Chemistry Laboratory Manual.profi maserNo ratings yet
- Kondawar 2017 Solvent Free Glycerol TransesterifiDocument11 pagesKondawar 2017 Solvent Free Glycerol TransesterifiElisabeta StamateNo ratings yet
- Cha 7Document13 pagesCha 7Tun Lin AungNo ratings yet
- Some Aspects of Calcium Phosphate Chemistry in Sugarcane ClarificationDocument8 pagesSome Aspects of Calcium Phosphate Chemistry in Sugarcane ClarificationJavier Manuel IbanezNo ratings yet
- Chemical Reactions of Copper LabDocument5 pagesChemical Reactions of Copper Labrikubean100% (1)
- Niosh 7303Document6 pagesNiosh 7303amaskpalsNo ratings yet
- Ikatan IonikDocument229 pagesIkatan IonikGani PurwiandonoNo ratings yet
- Gen. Chemistry 1: Quarter 1 - Module 2Document26 pagesGen. Chemistry 1: Quarter 1 - Module 2AniahsNelet80% (5)
- JEE Time Saver Course Planner FinalDocument24 pagesJEE Time Saver Course Planner FinalSadhak JainNo ratings yet
- Formulation and In-Vitro Characterization of Risperidone Nanosuspensions For The Enhancement of Drug Release RateDocument16 pagesFormulation and In-Vitro Characterization of Risperidone Nanosuspensions For The Enhancement of Drug Release RateSiva PrasadNo ratings yet
- Lec 1Document17 pagesLec 1Hadi H. HussenNo ratings yet
- Automated Blood Cell Analyzer (510712)Document34 pagesAutomated Blood Cell Analyzer (510712)ahsnupayaoNo ratings yet
- Topical Past PapersDocument208 pagesTopical Past PapersShafaq RamzanNo ratings yet
- Standard Costing & Variance AnalysisDocument23 pagesStandard Costing & Variance AnalysisZehra LeeNo ratings yet