You might also like
- Proceedings of The 16th Italian Association of Equine Veterinarians CongressDocument9 pagesProceedings of The 16th Italian Association of Equine Veterinarians CongressCabinet VeterinarNo ratings yet
- Zhao2019 - Invited Review Accelerating Mammary Gland InvolutionDocument17 pagesZhao2019 - Invited Review Accelerating Mammary Gland Involutionabdelaziz CHELIGHOUMNo ratings yet
- Proceedings of The 16th Italian Association of Equine Veterinarians CongressDocument6 pagesProceedings of The 16th Italian Association of Equine Veterinarians CongressCabinet VeterinarNo ratings yet
- 1 s2.0 S0093691X20303538 MainDocument11 pages1 s2.0 S0093691X20303538 MainRo BellingeriNo ratings yet
- Drug StudyDocument6 pagesDrug StudyJowel Cruz De LeonNo ratings yet
- Agalactia, Disgalactia y Nutrición de La Yegua Post PartoDocument6 pagesAgalactia, Disgalactia y Nutrición de La Yegua Post Partocarlo hernandezNo ratings yet
- The Myometrium: From Excitation To Contractions and Labour: Susan Wray and Clodagh PrendergastDocument31 pagesThe Myometrium: From Excitation To Contractions and Labour: Susan Wray and Clodagh PrendergastMBMNo ratings yet
- Daftar Pustaka 23Document7 pagesDaftar Pustaka 23sari idamihartiNo ratings yet
- Equine Placenta A Clinicians Perspective Part 1 Normal Placenta - Pozor - 2015Document8 pagesEquine Placenta A Clinicians Perspective Part 1 Normal Placenta - Pozor - 2015claudia igorNo ratings yet
- Daftar Pustaka 22Document6 pagesDaftar Pustaka 22sari idamihartiNo ratings yet
- LactationDocument8 pagesLactationDevuchandana RNo ratings yet
- MCN MidtermsDocument4 pagesMCN MidtermsMutya XDNo ratings yet
- M - Mask Adjustment R - Reposition Airway S - Suction Mouth and Nose O - Open Mouth P - Pressure Increase A - Airway AlternativeDocument4 pagesM - Mask Adjustment R - Reposition Airway S - Suction Mouth and Nose O - Open Mouth P - Pressure Increase A - Airway AlternativeSerious LeoNo ratings yet
- Eterinary Bstetrics: - Dr. S.BalasubramanianDocument413 pagesEterinary Bstetrics: - Dr. S.BalasubramanianprernaNo ratings yet
- Placenta Retention in The Cow: Report of Three Cases: Sokoto Journal of Veterinary SciencesDocument5 pagesPlacenta Retention in The Cow: Report of Three Cases: Sokoto Journal of Veterinary SciencesJuwita Tabeta DamayantiNo ratings yet
- Lactation PhysiologyDocument17 pagesLactation PhysiologysanathNo ratings yet
- Small Ruminant Neonatology 2008Document19 pagesSmall Ruminant Neonatology 2008daniellopesrpNo ratings yet
- Reproductive Physiology and Anatomy of The Sow PDFDocument4 pagesReproductive Physiology and Anatomy of The Sow PDFCarol OrequesNo ratings yet
- Embryo Transfer in The Dairy Herd: SuperovulationDocument4 pagesEmbryo Transfer in The Dairy Herd: SuperovulationVeteriner Hekim CengizhanNo ratings yet
- Physiology of Pregnancy & Functions of Placenta-1Document31 pagesPhysiology of Pregnancy & Functions of Placenta-1Ajit ShahNo ratings yet
- Contacciones UterinasDocument10 pagesContacciones UterinasEder RiveraNo ratings yet
- First Stage of Labor: Midwife Obstetrician InduceDocument2 pagesFirst Stage of Labor: Midwife Obstetrician InduceChe CacatianNo ratings yet
- Why Do We Perform Perineal Care?: DurationDocument5 pagesWhy Do We Perform Perineal Care?: DurationA CNo ratings yet
- 701-Article Text-2010-1-10-20221220Document6 pages701-Article Text-2010-1-10-20221220crokkapekanbaruNo ratings yet
- Canine Dystocia 38196 ArticleDocument6 pagesCanine Dystocia 38196 ArticlenurhasyiatnurNo ratings yet
- RAMOS-BN Drug Study Delivery RoomDocument2 pagesRAMOS-BN Drug Study Delivery RoomLouwella RamosNo ratings yet
- Akers 1990Document16 pagesAkers 1990ccamachoNo ratings yet
- Ontogeny of Human Pancreatic Exocrine Function: Regular ReviewDocument4 pagesOntogeny of Human Pancreatic Exocrine Function: Regular ReviewFernandaNo ratings yet
- Uterine Involution & Postpartum Ovarian ActivityDocument8 pagesUterine Involution & Postpartum Ovarian Activityjerry smatt dumtoNo ratings yet
- MCN Notes 2Document26 pagesMCN Notes 2Rogeimar Claire MangomayaoNo ratings yet
- Task IncomingDocument14 pagesTask Incomingmonica mittiamNo ratings yet
- Milk Somatic Cells, Factors Influencing Their Release, Future ProspectsDocument16 pagesMilk Somatic Cells, Factors Influencing Their Release, Future Prospectsจักรพงศ์ กิ่วแก้วNo ratings yet
- Clinical Benefits of Lactoferrin For Infants and Children: Paolo Manzoni, MD, PHDDocument10 pagesClinical Benefits of Lactoferrin For Infants and Children: Paolo Manzoni, MD, PHDRenata HoneyNo ratings yet
- OXITOCINDocument7 pagesOXITOCINhoneyNo ratings yet
- Generic NameDocument3 pagesGeneric NameSittie Nashieva A. UsmanNo ratings yet
- OB HesiDocument3 pagesOB HesiLeena Lapena100% (4)
- Chapter 9: Nursing Care During Normal Pregnancy and Care of The Developing FetusDocument8 pagesChapter 9: Nursing Care During Normal Pregnancy and Care of The Developing FetusAlyssaGrandeMontimor100% (1)
- NS 1 Quiz 3Document7 pagesNS 1 Quiz 3Marah PacioNo ratings yet
- Oxytocin Drug StudyDocument3 pagesOxytocin Drug Studysachi yuNo ratings yet
- MCN ReportDocument21 pagesMCN ReportJœnríčk AzueloNo ratings yet
- Induction of LabourDocument51 pagesInduction of LabourSarita PariyarNo ratings yet
- B55. PPH - TechUpdate2Document2 pagesB55. PPH - TechUpdate2jhon heriansyahNo ratings yet
- Material ON Drugs Used IN ObstetricsDocument5 pagesMaterial ON Drugs Used IN ObstetricsMallika JoonNo ratings yet
- Revised-Case-based - Scenario - Day - BANZON, HAZEL ANNE D.Document4 pagesRevised-Case-based - Scenario - Day - BANZON, HAZEL ANNE D.Hazel Anne BanzonNo ratings yet
- Nursing Care PlansDocument4 pagesNursing Care PlansRITIK KUMARNo ratings yet
- K11 12 Fisiologi KehamilanDocument75 pagesK11 12 Fisiologi KehamilanAditya PrabawaNo ratings yet
- Management of Hyperlactation Syndrome by Full DraiDocument3 pagesManagement of Hyperlactation Syndrome by Full Draik_klinoxNo ratings yet
- (1) Manejo e fertilida de éguas criadas com sêmen congeladoDocument10 pages(1) Manejo e fertilida de éguas criadas com sêmen congeladoIsabellaNo ratings yet
- Drug Study OF Medications Used in Delivery Room: Republic of The Philippines Mindanao State University Marawi CityDocument19 pagesDrug Study OF Medications Used in Delivery Room: Republic of The Philippines Mindanao State University Marawi CityHoney Semafranca PlatolonNo ratings yet
- Effective uterotonics for postpartum hemorrhage preventionDocument13 pagesEffective uterotonics for postpartum hemorrhage preventionNicole Andrea Silva GaeteNo ratings yet
- Age of Weaning Lambs: A Greener World Technical Advice Fact Sheet No. 17Document7 pagesAge of Weaning Lambs: A Greener World Technical Advice Fact Sheet No. 17Fatmalina AliNo ratings yet
- Phy Ist Stag LBRDocument9 pagesPhy Ist Stag LBRAparna KinginiNo ratings yet
- 2011 Introduction To Maternity Pediatric Nursing Solution Manual 6th EditionDocument24 pages2011 Introduction To Maternity Pediatric Nursing Solution Manual 6th EditionAndrewFoleydegk100% (42)
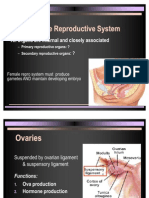
- Female Repro System Must Produce Gametes AND Maintain Developing EmbryoDocument27 pagesFemale Repro System Must Produce Gametes AND Maintain Developing EmbryoAbhishek GuptaNo ratings yet
- Ncma217 Week 11Document4 pagesNcma217 Week 11Polly ArcheronNo ratings yet
- Lecture N4 Breast-Feeding. Value and Advantages of Breast-Feeding. Methods of Calculation of Daily Volume of Feeding. Weaning at Breast FeedingDocument7 pagesLecture N4 Breast-Feeding. Value and Advantages of Breast-Feeding. Methods of Calculation of Daily Volume of Feeding. Weaning at Breast FeedingAVINASH PvkNo ratings yet
- BSN-II Labor InterventionsDocument3 pagesBSN-II Labor InterventionsDina Jane BernalNo ratings yet
- Retained Placenta GuidelinesDocument3 pagesRetained Placenta GuidelineschaeworldNo ratings yet
- 1.1c Implantation & Placental DevelopmentDocument5 pages1.1c Implantation & Placental DevelopmentBea Samonte100% (1)
- Study On Lab AnimalDocument5 pagesStudy On Lab AnimalKishan PatelNo ratings yet
- A Case Report On Wound Healing Activity of Cow GheeDocument4 pagesA Case Report On Wound Healing Activity of Cow GheeKishan PatelNo ratings yet
- Evaluation of Wisprec Advanced Spray and Certain Phytopharmaceutical Prototypes For Their Topical Anti-Inflammatory EffectDocument6 pagesEvaluation of Wisprec Advanced Spray and Certain Phytopharmaceutical Prototypes For Their Topical Anti-Inflammatory EffectKishan PatelNo ratings yet
- 1 s2.0 S0022030298758205 MainDocument10 pages1 s2.0 S0022030298758205 MainKishan PatelNo ratings yet
- A 100-Year Review: Stress Physiology Including Heat Stress: J. Dairy Sci. 100:10367-10380Document14 pagesA 100-Year Review: Stress Physiology Including Heat Stress: J. Dairy Sci. 100:10367-10380Kishan PatelNo ratings yet
- A 100-Year Review: Stress Physiology Including Heat Stress: J. Dairy Sci. 100:10367-10380Document14 pagesA 100-Year Review: Stress Physiology Including Heat Stress: J. Dairy Sci. 100:10367-10380Kishan PatelNo ratings yet
- (24537837 - Folia Veterinaria) Types of Wounds and The Prevalence of Bacterial Contamination of Wounds in The Clinical Practice of Small AnimalsDocument9 pages(24537837 - Folia Veterinaria) Types of Wounds and The Prevalence of Bacterial Contamination of Wounds in The Clinical Practice of Small AnimalsKishan PatelNo ratings yet
- A Review On Mastitis: International Journal of Medicine ResearchDocument6 pagesA Review On Mastitis: International Journal of Medicine ResearchKishan PatelNo ratings yet
- A Review On Mastitis: International Journal of Medicine ResearchDocument6 pagesA Review On Mastitis: International Journal of Medicine ResearchKishan PatelNo ratings yet
- Study On Lab AnimalDocument5 pagesStudy On Lab AnimalKishan PatelNo ratings yet
- Efficacy of Styplon Vet Bolus for Hemorrhagic ConditionsDocument2 pagesEfficacy of Styplon Vet Bolus for Hemorrhagic ConditionsKishan PatelNo ratings yet
- Nutrition and Wound Healing: Content CreatorsDocument50 pagesNutrition and Wound Healing: Content CreatorsKishan PatelNo ratings yet
- Effect - of - Probiotics - Supplementation - On - Milk - Yield - Biobloom Zydus Product (Similar Article)Document6 pagesEffect - of - Probiotics - Supplementation - On - Milk - Yield - Biobloom Zydus Product (Similar Article)Kishan PatelNo ratings yet
- MSD - IMP (Controversial Use of Vitamin A)Document1 pageMSD - IMP (Controversial Use of Vitamin A)Kishan PatelNo ratings yet
- Commented (d1) : Can Use Any One of The Tag LineDocument3 pagesCommented (d1) : Can Use Any One of The Tag LineKishan PatelNo ratings yet
- LCNCDocument14 pagesLCNCDuniel OrtuñoNo ratings yet
- Micro Organisms ReviewerDocument72 pagesMicro Organisms ReviewerdavenNo ratings yet
- Silkworm moth: from egg to emergenceDocument78 pagesSilkworm moth: from egg to emergenceSoubhik MondalNo ratings yet
- Pathology For The Health Professions 5th Edition Damjanov Solutions ManualDocument25 pagesPathology For The Health Professions 5th Edition Damjanov Solutions ManualKevinRobertsbfak98% (56)
- WBCs Chap 2Document119 pagesWBCs Chap 2Hoor ul Ain RamzanNo ratings yet
- Vertical Root FractureDocument2 pagesVertical Root FractureJeremiah SchmidtNo ratings yet
- Soalan Objektif Mac2014Document2 pagesSoalan Objektif Mac2014Siti NajwaNo ratings yet
- Obstructive Disease of Lung and Physiotherapy ManagementDocument76 pagesObstructive Disease of Lung and Physiotherapy Managementphysio43100% (1)
- Basic Science JSS2 Question BankDocument3 pagesBasic Science JSS2 Question BankIsaacNo ratings yet
- Mutationbreedingppt 161203184149Document41 pagesMutationbreedingppt 161203184149MuneebNo ratings yet
- Sodium Stibogluconate Pentamidine IsetionateDocument10 pagesSodium Stibogluconate Pentamidine IsetionateThunder BoltNo ratings yet
- Archive of SID: Riginal RticleDocument4 pagesArchive of SID: Riginal RticleDyah Putri Ayu DinastyarNo ratings yet
- EMT Chapter 10 Vocabulary Flashcards - PT 1Document4 pagesEMT Chapter 10 Vocabulary Flashcards - PT 1Emily CallerNo ratings yet
- Ambroxol Prescribing InformationDocument6 pagesAmbroxol Prescribing InformationManmohan SinghNo ratings yet
- PHYSIO ReviewerDocument3 pagesPHYSIO ReviewerPHILYP EPHRAIM PARANGALANNo ratings yet
- Introduction To Blood BankingDocument3 pagesIntroduction To Blood Bankingsam hermosadoNo ratings yet
- IAS Biology Student Book 1 (2018) AnswersDocument74 pagesIAS Biology Student Book 1 (2018) AnswersGazar76% (33)
- BotanistsDocument2 pagesBotanistsRika MuhiNo ratings yet
- Major Basic Set (Operating Room Instruments)Document5 pagesMajor Basic Set (Operating Room Instruments)Ralph Jetzon MorillaNo ratings yet
- You Have A BrainDocument28 pagesYou Have A BrainZondervan91% (11)
- 2022 Anatmoy and Physiology Cheat Sheet!Document3 pages2022 Anatmoy and Physiology Cheat Sheet!Estella BrockwellNo ratings yet
- Anatomyand Physiologyofthe Thoraciclymphatic SystemDocument15 pagesAnatomyand Physiologyofthe Thoraciclymphatic SystemLis Borda MuñozNo ratings yet
- Meat and Fish Food SciencesDocument19 pagesMeat and Fish Food SciencesAbeer Basharat100% (1)
- FluidDocument190 pagesFluidAndrias OzNo ratings yet
- PacemakersDocument6 pagesPacemakersFirst LastNo ratings yet
- Practice Midterm ReviewDocument14 pagesPractice Midterm Reviewasjiodjw100% (2)
- Biofact Sheet Nerves SynapsesDocument4 pagesBiofact Sheet Nerves SynapsesJonMortNo ratings yet
- Lesson Plan in Arteries and VeinsDocument3 pagesLesson Plan in Arteries and VeinsHarold BadillaNo ratings yet
- What Makes It Beautiful VitaDocument8 pagesWhat Makes It Beautiful VitaVladimir DrăgoiNo ratings yet
- Blood Pressure in Non-Critically Ill Preterm and Full-Term NeonatesDocument9 pagesBlood Pressure in Non-Critically Ill Preterm and Full-Term NeonatesEduardo Rios DuboisNo ratings yet