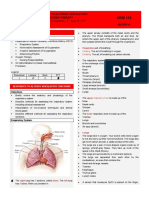
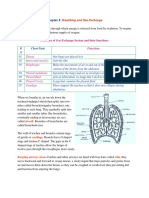
You might also like
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (589)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (842)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5806)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- DR Breath by Carl Stough PDFDocument128 pagesDR Breath by Carl Stough PDFJuan Vizán86% (7)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Eukaryotic DNA ReplicationDocument25 pagesEukaryotic DNA ReplicationKimberly Thomas100% (1)
- Year 8 Science Progress 3 Practice Questions and AnswersDocument23 pagesYear 8 Science Progress 3 Practice Questions and AnswersMohammed100% (1)
- D - 5answer KeyDocument15 pagesD - 5answer KeyJune DumdumayaNo ratings yet
- Nursing Care of The High Risk Newborn To Maturity: Nursing Diagnosis Outcome Evaluation Assessment InterventionDocument9 pagesNursing Care of The High Risk Newborn To Maturity: Nursing Diagnosis Outcome Evaluation Assessment InterventionNur Sanaani100% (1)
- NCP-Ineffective AirwayDocument5 pagesNCP-Ineffective Airwayjava_biscocho1229No ratings yet
- 118 Skills Lab-Week 1-Responses To Altered Ventilatory FunctionsDocument8 pages118 Skills Lab-Week 1-Responses To Altered Ventilatory FunctionsKeisha BartolataNo ratings yet
- 4 A Lesson PLan RevisedDocument7 pages4 A Lesson PLan RevisedChrismarie Mae PanoyNo ratings yet
- Causative Agents Causative Agents: Headache, Myalgia, & Nausea Are Added S/SX For Streptococcal PharyngitisDocument21 pagesCausative Agents Causative Agents: Headache, Myalgia, & Nausea Are Added S/SX For Streptococcal PharyngitisDon Chiaw Manongdo100% (1)
- The Cytoskeleton #2: Molecular MotorsDocument34 pagesThe Cytoskeleton #2: Molecular MotorsKimberly ThomasNo ratings yet
- The Cytoskeleton #1: Cytoskeletal FilamentsDocument42 pagesThe Cytoskeleton #1: Cytoskeletal FilamentsKimberly ThomasNo ratings yet
- Lab 2 - Biochemical Identification of Unknown BacteriaDocument9 pagesLab 2 - Biochemical Identification of Unknown BacteriaKimberly ThomasNo ratings yet
- LAB 6 - HintsDocument2 pagesLAB 6 - HintsKimberly ThomasNo ratings yet
- Unit - I APCDocument37 pagesUnit - I APCMani DeepNo ratings yet
- Throat Mcqs Points Rida NaqviDocument5 pagesThroat Mcqs Points Rida NaqviÅli Raza ChaudaryNo ratings yet
- The Skin Is The Largest Organ in Your BodyDocument4 pagesThe Skin Is The Largest Organ in Your BodyLV BENDANANo ratings yet
- Quiz Cell 3oktDocument25 pagesQuiz Cell 3oktFarah AsnidaNo ratings yet
- Nayli Bio SBP 2019 EseiDocument5 pagesNayli Bio SBP 2019 EseiEqis AlfiesyaNo ratings yet
- Pediatric Chest PhysiotherapyDocument25 pagesPediatric Chest PhysiotherapyShubha DiwakarNo ratings yet
- Mechanical VentilationDocument12 pagesMechanical VentilationDoru VoicuNo ratings yet
- Respiratory SystemDocument89 pagesRespiratory SystemMuhammad HanifNo ratings yet
- 2.1 (I) Know The Properties of Gas Exchange Surfaces in Living Organisms 2Document16 pages2.1 (I) Know The Properties of Gas Exchange Surfaces in Living Organisms 2YEUNG WAN CHUEN ANGELIQUENo ratings yet
- LIFE PROCESSES - Question BankDocument15 pagesLIFE PROCESSES - Question BankMISHKA KHANDELWALNo ratings yet
- 1 - Normal Saline For Endotracheal SuctioningDocument28 pages1 - Normal Saline For Endotracheal SuctioningRena WidyaNo ratings yet
- 11 Lung PathologyDocument7 pages11 Lung PathologycarlgangcaNo ratings yet
- JUE-410F Installation Manual PDFDocument74 pagesJUE-410F Installation Manual PDFВячеславNo ratings yet
- Efficacy of Pursed-Lips Breathing: A Breathing Pattern Retraining Strategy For Dyspnea ReductionDocument8 pagesEfficacy of Pursed-Lips Breathing: A Breathing Pattern Retraining Strategy For Dyspnea ReductionGaoudam NatarajanNo ratings yet
- BPL Multi para Monitor Excello - Excello-D Excello-MDocument4 pagesBPL Multi para Monitor Excello - Excello-D Excello-MSujeet MurmuNo ratings yet
- Breathing and Gas Exchange: RespirationDocument4 pagesBreathing and Gas Exchange: RespirationAbubakar Siddiq RaminNo ratings yet
- Worksheet Respiratory SystemDocument2 pagesWorksheet Respiratory SystemAnonymous piAtTTKkNo ratings yet
- Activity For Set B - 08302022Document1 pageActivity For Set B - 08302022ANALYN REYESNo ratings yet
- CARE Dept. of Interventional Pulmonology - 3Document7 pagesCARE Dept. of Interventional Pulmonology - 3rohit6varma-4No ratings yet
- O-Integrated Science Syllabus PDFDocument36 pagesO-Integrated Science Syllabus PDFBrighton Mhlanga89% (9)
- Oxygen Therapy Learning Module Cat 1and2Document62 pagesOxygen Therapy Learning Module Cat 1and2Baha'aeddin HammadNo ratings yet
- 2014 The ANZICS Percutaneous Dilatational Tracheostomy Consensus StatementDocument25 pages2014 The ANZICS Percutaneous Dilatational Tracheostomy Consensus StatementyudiNo ratings yet