You might also like
- Silicones&Sulfates - Smart Usage Manual - (2023)Document28 pagesSilicones&Sulfates - Smart Usage Manual - (2023)Alifia AzhaaraNo ratings yet
- Lyphochek Immunoassay Plus Control Levels 1, 2 and 3Document11 pagesLyphochek Immunoassay Plus Control Levels 1, 2 and 3Enrique DuarteNo ratings yet
- AppendixDocument11 pagesAppendixapi-3800038No ratings yet
- Alkynyl-Functionalized Gold NHC Complexes and TheiDocument5 pagesAlkynyl-Functionalized Gold NHC Complexes and TheikarthickrajaNo ratings yet
- Articulo Numero 1Document5 pagesArticulo Numero 1Andrés OrtizNo ratings yet
- Colombo1999 PDFDocument5 pagesColombo1999 PDFJulianDavidMorenoNo ratings yet
- Study of A Single Pass Shell and Tube Heat ExchangerDocument21 pagesStudy of A Single Pass Shell and Tube Heat Exchangermahbub1332No ratings yet
- Supp Data 6 JDP 271115 PDFDocument28 pagesSupp Data 6 JDP 271115 PDFSascha KoenenNo ratings yet
- 1.2003viscosities, Densities, and Ultrasonic Velocities of 3-Pentanone +Document4 pages1.2003viscosities, Densities, and Ultrasonic Velocities of 3-Pentanone +Sanigdha AcharyaNo ratings yet
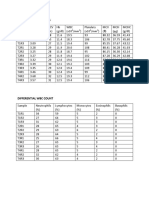
- Variable 1 Variable 2: Computed ValueDocument13 pagesVariable 1 Variable 2: Computed ValueEONSNo ratings yet
- Chaos in Fractional-Order Autonomous Nonlinear Systems: Wajdi M. Ahmad, J.C. SprottDocument13 pagesChaos in Fractional-Order Autonomous Nonlinear Systems: Wajdi M. Ahmad, J.C. SprottAlfredo Soliz GamboaNo ratings yet
- Hujan Rancangan Analitis Kala Ulang 100 TahunDocument24 pagesHujan Rancangan Analitis Kala Ulang 100 TahunArthur Aditya YohanisNo ratings yet
- The University of Jordan: Omar Thabet Ahmad Alqasser Mahmoud Abu RamadanDocument13 pagesThe University of Jordan: Omar Thabet Ahmad Alqasser Mahmoud Abu RamadanEssa HabashNo ratings yet
- Water Quality of The Tazerbo Wellfield FDocument6 pagesWater Quality of The Tazerbo Wellfield FfarajelmabroukNo ratings yet
- Woodman 1925Document4 pagesWoodman 1925edmentzNo ratings yet
- DNA Typing of Short Tandem Repeat Loci On Y-Chr of Greeh PopulationDocument4 pagesDNA Typing of Short Tandem Repeat Loci On Y-Chr of Greeh PopulationThanasis PapatheodorouNo ratings yet
- (2755205X) Assessed ProblemsDocument12 pages(2755205X) Assessed Problems肖宇翔No ratings yet
- Power System Analysis - Stevenson - Grainger - AnnexDocument5 pagesPower System Analysis - Stevenson - Grainger - AnnexSaghir AhmadNo ratings yet
- 244 Experiment 1 (1) - 1Document7 pages244 Experiment 1 (1) - 1Bophepa MaseletsaneNo ratings yet
- Solubility of Carbon Dioxide in Aqueous Solutions of Sodium Chloride: Experimental Results and CorrelationDocument18 pagesSolubility of Carbon Dioxide in Aqueous Solutions of Sodium Chloride: Experimental Results and CorrelationzibaNo ratings yet
- Experiment 1Document6 pagesExperiment 1Botlhe Kgotla SamNo ratings yet
- Lampiran 4. Analisis Data Limfosit Ikan Mas (Cyprinus Carpio L.) Pada Toksisitas Subletal Limbah Cair Batik Hasil BiosorpsiDocument6 pagesLampiran 4. Analisis Data Limfosit Ikan Mas (Cyprinus Carpio L.) Pada Toksisitas Subletal Limbah Cair Batik Hasil BiosorpsiHana AgustianaNo ratings yet
- Condenser DesignDocument25 pagesCondenser DesignmohsinziaNo ratings yet
- Petroleum Engineering Lab 1 (Pge313) Experiment 3 Determination of PermeabilityDocument20 pagesPetroleum Engineering Lab 1 (Pge313) Experiment 3 Determination of PermeabilityAli AbubakarNo ratings yet
- Exp 8 Lab ReportDocument7 pagesExp 8 Lab ReportSEHLACWINo ratings yet
- HypothesisDocument20 pagesHypothesisbbbbNo ratings yet
- 0010 - El-Hammamy 2011Document5 pages0010 - El-Hammamy 2011Amr GamalNo ratings yet
- TC NTC 1 DatasheetDocument4 pagesTC NTC 1 DatasheetIvan MillánNo ratings yet
- Crystal Structure of t-boc-O-benzyl-L-tyrosyl-D-alanyl-L - (O-Benzyl) - Glutamate: MonohydrateDocument8 pagesCrystal Structure of t-boc-O-benzyl-L-tyrosyl-D-alanyl-L - (O-Benzyl) - Glutamate: MonohydrateNaina MarbusNo ratings yet
- Data Repository MaterialDocument31 pagesData Repository MaterialMarcio AlbuquerqueNo ratings yet
- Ce 261 PS4 PDFDocument5 pagesCe 261 PS4 PDFFerdz PequiroNo ratings yet
- Rules of Thumb - AviationDocument26 pagesRules of Thumb - AviationMartin Cernic100% (1)
- Viscosity of c5h802 - 28Document1 pageViscosity of c5h802 - 28CharlesNo ratings yet
- IEM-19-29 M.Suleman (DOE) A6Document11 pagesIEM-19-29 M.Suleman (DOE) A6workassassinNo ratings yet
- Esther BloodDocument2 pagesEsther BloodAnietie AnsaNo ratings yet
- Scleroderma ExperimentsDocument5 pagesScleroderma ExperimentsspartangyalonlyNo ratings yet
- Linear Statistical ModelsDocument16 pagesLinear Statistical ModelssdeeproyNo ratings yet
- PDP, Hyd Microsatellite Variability and Their Association With Growth and Egg Production Traits in ChickenDocument22 pagesPDP, Hyd Microsatellite Variability and Their Association With Growth and Egg Production Traits in Chickenvignesh murugesanNo ratings yet
- Jo9b02466 Si 001Document44 pagesJo9b02466 Si 001Oscaro TorresNo ratings yet
- Database XRDDocument6 pagesDatabase XRDwindaNo ratings yet
- Gmrp-Libya FinalDocument22 pagesGmrp-Libya FinalfarajelmabroukNo ratings yet
- Oxygen Isotope Calibration of The Onset of Ice-RaftingDocument4 pagesOxygen Isotope Calibration of The Onset of Ice-Rafting7hxgxzkzqnNo ratings yet
- Supporting InformationDocument18 pagesSupporting Informationding.hanbo0219No ratings yet
- Notes 109Document26 pagesNotes 109RAISHANo ratings yet
- Isobaric Vapor-Liquid and Liquid-Liquid Equilibria For Chloroform + Methanol + 1-Ethyl-3-Methylimidazolium Trifluoromethanesulfonate at 100 KpaDocument6 pagesIsobaric Vapor-Liquid and Liquid-Liquid Equilibria For Chloroform + Methanol + 1-Ethyl-3-Methylimidazolium Trifluoromethanesulfonate at 100 Kpaluis fernando muro cordovaNo ratings yet
- NO. Elements Page NoDocument21 pagesNO. Elements Page NoandriasNo ratings yet
- Supplementary Data For Genetic Study of Ancient CanaanitesDocument23 pagesSupplementary Data For Genetic Study of Ancient CanaanitesGirawa KandaNo ratings yet
- Explore: Case Processing SummaryDocument12 pagesExplore: Case Processing SummaryPrajoko SusiloNo ratings yet
- Bookmarks PDFDocument17 pagesBookmarks PDFAniket DubeyNo ratings yet
- Variable 1 Variable 2Document4 pagesVariable 1 Variable 2Jayleeteukie TyelfNo ratings yet
- Nichrome - WikipediaDocument12 pagesNichrome - Wikipediamei krismahariyantoNo ratings yet
- SI JACS CHB PTDocument84 pagesSI JACS CHB PT192.22irNo ratings yet
- Harrison2013 Article IsolationAndCharacterizationOf PrimersDocument3 pagesHarrison2013 Article IsolationAndCharacterizationOf PrimersYasmin Gálvez MuñozNo ratings yet
- University of Santo Tomas CHE 514L: Chemical Engineering Laboratory IIDocument6 pagesUniversity of Santo Tomas CHE 514L: Chemical Engineering Laboratory IIKelly Sheine SisonNo ratings yet
- Equation of State of Brucite: Single-Crystal Brillouin Spectroscopy Study and Polycrystalline Pressure-Volume-Temperature MeasurementDocument7 pagesEquation of State of Brucite: Single-Crystal Brillouin Spectroscopy Study and Polycrystalline Pressure-Volume-Temperature Measurementmauricio rojas alvarezNo ratings yet
- Nama Alber Erfan Mohammad No. BP 1610413003Document12 pagesNama Alber Erfan Mohammad No. BP 1610413003studynyetNo ratings yet
- Duran Valencia2001Document4 pagesDuran Valencia2001jasontodd22031995No ratings yet
- InfoDocument9 pagesInfoMadhu DuraichamyNo ratings yet
- Andalan - Lab No.2Document4 pagesAndalan - Lab No.2Jolo AndalanNo ratings yet
- An Introduction to Gas Discharges: Pergamon International Library of Science, Technology, Engineering and Social StudiesFrom EverandAn Introduction to Gas Discharges: Pergamon International Library of Science, Technology, Engineering and Social StudiesNo ratings yet
- 1.4 Pharmacy As My Future SpecialtyDocument3 pages1.4 Pharmacy As My Future SpecialtyDanti Nafis100% (1)
- Class-X Science Pre Board Q.p-2020-21FinalDocument12 pagesClass-X Science Pre Board Q.p-2020-21FinalRoshan Kumar SahooNo ratings yet
- 9.FT ProcedureDocument10 pages9.FT ProceduremadanbhopeNo ratings yet
- Selina Solutions Concise Chemistry For Class 10 Chapter 9Document3 pagesSelina Solutions Concise Chemistry For Class 10 Chapter 9Naved Shamsi 78No ratings yet
- Periodic Classification - DPP 2 (Of Lec 03) - Arjuna JEE 2.0 2024Document2 pagesPeriodic Classification - DPP 2 (Of Lec 03) - Arjuna JEE 2.0 2024weeb.with.internet.accessNo ratings yet
- Elementis Bentone 27 TdsDocument2 pagesElementis Bentone 27 Tdsmgamal1080No ratings yet
- Tuberculosis EssayDocument4 pagesTuberculosis Essaybmakkoaeg100% (3)
- PT11Document18 pagesPT11Pradeep Kumar BowmarajuNo ratings yet
- The Election of Andrew Jackson To The Presidency in 1928 Marked The Political Ascendancy of TheDocument50 pagesThe Election of Andrew Jackson To The Presidency in 1928 Marked The Political Ascendancy of TheFREESHIP AsistantNo ratings yet
- MA8110 TDS Rev11Document2 pagesMA8110 TDS Rev11Rushikesh DeshpandeNo ratings yet
- CMS177-Nicotinic Acid (Vit B3)Document2 pagesCMS177-Nicotinic Acid (Vit B3)darrendelfinoy9No ratings yet
- gmr18510 - Simple and Inexpensive Modified Ctab and Silica Particles Based Method Recalcitrant PlantDocument9 pagesgmr18510 - Simple and Inexpensive Modified Ctab and Silica Particles Based Method Recalcitrant Plantlouisehip UFCNo ratings yet
- POLYCARBONEDocument11 pagesPOLYCARBONEAshley Noemi Sanchez DelgadoNo ratings yet
- Poro MaxDocument9 pagesPoro MaxJohanes GigihNo ratings yet
- Swing Check Valve Rubber DiscDocument2 pagesSwing Check Valve Rubber DiscsathishNo ratings yet
- Hydrodynamic LubricationDocument9 pagesHydrodynamic LubricationLakshitha Akalanka VithanageNo ratings yet
- EVS MapsDocument18 pagesEVS MapsGaurav GiriNo ratings yet
- 11 Chemistry Notes Ch04 Chemical Bonding and Molecular StructureDocument25 pages11 Chemistry Notes Ch04 Chemical Bonding and Molecular Structureshah khisarwNo ratings yet
- Ipal - Modul 2 - Analisis Karakteristik Air LimbahDocument7 pagesIpal - Modul 2 - Analisis Karakteristik Air LimbahAndhika HerdiawanNo ratings yet
- Colligative Properties of Solutions ReviewerDocument4 pagesColligative Properties of Solutions ReviewerPrecious Lara MangobaNo ratings yet
- ECH 3903 Lab ManualDocument103 pagesECH 3903 Lab ManualDivagar SivaselvamNo ratings yet
- Chemistry 112 Dye Kinetics Laboratory 2011Document4 pagesChemistry 112 Dye Kinetics Laboratory 2011Lavenia Alou MagnoNo ratings yet
- Hexion Vodnye Otverditeli I Smoly EngDocument6 pagesHexion Vodnye Otverditeli I Smoly EngCloudy DayNo ratings yet
- Chemistry Module Iii Inorganic Chemistry For Iit Jee Main and Advanced Vineet Agarwal Mcgraw Hill Education Vineet Agarwal Full Chapter PDF ScribdDocument68 pagesChemistry Module Iii Inorganic Chemistry For Iit Jee Main and Advanced Vineet Agarwal Mcgraw Hill Education Vineet Agarwal Full Chapter PDF Scribdeula.gibbs793100% (4)
- NeoCryl XK-205 PdsDocument1 pageNeoCryl XK-205 PdsLeandro EsvizaNo ratings yet
- Syllabus in Chemical Biology III (Biomolecules) (TCB3)Document12 pagesSyllabus in Chemical Biology III (Biomolecules) (TCB3)Yuri PaderesNo ratings yet
- SLG Chem3 LG 2.15 Application of HalogenoalkanesDocument4 pagesSLG Chem3 LG 2.15 Application of HalogenoalkanesLorraine CalacsanNo ratings yet
- Chapter 22: Introduction To Electroanalytical ChemistryDocument29 pagesChapter 22: Introduction To Electroanalytical ChemistryMedina KimJunelNo ratings yet
- As Physics P2 Topical Part IDocument390 pagesAs Physics P2 Topical Part Iabdyllah22.nomanNo ratings yet