You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5796)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Disordersofrumen Distensionanddysmotility: Derek FosterDocument14 pagesDisordersofrumen Distensionanddysmotility: Derek FosterSara ConnorNo ratings yet
- Vagus Indigestion in Bovines: A Review in Historical PerspectiveDocument8 pagesVagus Indigestion in Bovines: A Review in Historical PerspectiveSara ConnorNo ratings yet
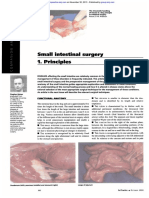
- Sall Intestinal Surgery Techniques: Cllipi) TicallDocument14 pagesSall Intestinal Surgery Techniques: Cllipi) TicallSara ConnorNo ratings yet
- In Practice-2012-Hayes-250-9Document9 pagesIn Practice-2012-Hayes-250-9Sara ConnorNo ratings yet
- MacFarlane - Chronic Pain in DogsDocument9 pagesMacFarlane - Chronic Pain in DogsSara ConnorNo ratings yet
- In Practice-2000-Baines-502-17Document12 pagesIn Practice-2000-Baines-502-17Sara ConnorNo ratings yet
- 9 - Pdfsam - Vsu12006 - Assessment of Surgical SkillsDocument1 page9 - Pdfsam - Vsu12006 - Assessment of Surgical SkillsSara ConnorNo ratings yet
- Suture Materials and Patterns PDFDocument9 pagesSuture Materials and Patterns PDFSara ConnorNo ratings yet
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (589)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Chapter 056Document42 pagesChapter 056Adistri KimNo ratings yet
- Liquichek Elevated CRP Control Levels 1, 2 and 3: Revision Date 2022-05-26 Indicates Revised InformationDocument1 pageLiquichek Elevated CRP Control Levels 1, 2 and 3: Revision Date 2022-05-26 Indicates Revised InformationowoladeidowuNo ratings yet
- Absorption & AssimilationDocument16 pagesAbsorption & AssimilationNaan Mahan AlleNo ratings yet
- Digestive SystemDocument7 pagesDigestive SystemEva BrendaNo ratings yet
- FASTADocument3 pagesFASTAM.Akram TatriNo ratings yet
- Index 2019 Recent Developments in Applied Microbiology and BiochemistryDocument14 pagesIndex 2019 Recent Developments in Applied Microbiology and BiochemistryramamurthiNo ratings yet
- FullDocument350 pagesFullgeorgeNo ratings yet
- Polyplus Transfection® Life Science Research and Non-Viral In-Vivo Transfection ReagentsDocument6 pagesPolyplus Transfection® Life Science Research and Non-Viral In-Vivo Transfection ReagentsDiego GarridoNo ratings yet
- Inmed Animal Health - XLSX SheetDocument27 pagesInmed Animal Health - XLSX SheetInmed AnimalNo ratings yet
- Cross interactions between the Alzheimer's disease amyloid-β peptide and other amyloid proteins: a further aspect of the amyloid cascade hypothesisDocument17 pagesCross interactions between the Alzheimer's disease amyloid-β peptide and other amyloid proteins: a further aspect of the amyloid cascade hypothesisMAYRA PAZNo ratings yet
- Trial Sem 1 2017 SMK THREE RIVERS Answers PDFDocument12 pagesTrial Sem 1 2017 SMK THREE RIVERS Answers PDFChee Beng YeapNo ratings yet
- Biochem Lab QuipperDocument4 pagesBiochem Lab QuipperJULIANNE BAYHONNo ratings yet
- The Digestive SystemDocument5 pagesThe Digestive SystemFarheen AmerNo ratings yet
- NEWS 1306 US TitlelistDocument76 pagesNEWS 1306 US TitlelistKartheek RajaNo ratings yet
- Nutrition and Diet Therapy: Section 1 Chapter 1: The Relationship of Food and Health Nutrients VitaminsDocument32 pagesNutrition and Diet Therapy: Section 1 Chapter 1: The Relationship of Food and Health Nutrients VitaminsAnonymousTargetNo ratings yet
- Proteinsn and Nucleic AcidsDocument17 pagesProteinsn and Nucleic AcidsJasmin RIshel Dela CruzNo ratings yet
- Quiz #3: Biochemical Engineering Fall 2003Document5 pagesQuiz #3: Biochemical Engineering Fall 2003Princess Janine CatralNo ratings yet
- Biochemistry Unit ConversionDocument4 pagesBiochemistry Unit ConversionTran Trong NghiaNo ratings yet
- DNA - and RNA-Based Computing SystemsDocument397 pagesDNA - and RNA-Based Computing SystemsAgustin BarrientosNo ratings yet
- (Methods in Molecular Biology 1644) Vladimir V. Didenko (Eds.) - Fast Detection of DNA Damage - Methods and Protocols-Springer New York (2017) PDFDocument213 pages(Methods in Molecular Biology 1644) Vladimir V. Didenko (Eds.) - Fast Detection of DNA Damage - Methods and Protocols-Springer New York (2017) PDFMauro Porcel de PeraltaNo ratings yet
- Keywords: Protease, Lipase, Visceral Enzyme, Mackerel FishDocument9 pagesKeywords: Protease, Lipase, Visceral Enzyme, Mackerel FishlidyawatiNo ratings yet
- Exercise 13Document27 pagesExercise 13Jamie Paola SevillaNo ratings yet
- Nuceic Acid ProjectDocument2 pagesNuceic Acid ProjectCaryl Alvarado SilangNo ratings yet
- Varun II SeminarDocument15 pagesVarun II Seminarchidananda sNo ratings yet
- EnzymeDocument31 pagesEnzymemiriam harriottNo ratings yet
- QuinoaFacts OthersDocument186 pagesQuinoaFacts OthersBanSnake_No ratings yet
- A Novel Sample Preparation For Shotgun Proteomics Characterization of HCPs in AntibodiesDocument9 pagesA Novel Sample Preparation For Shotgun Proteomics Characterization of HCPs in AntibodiesTaylor SimmonnsNo ratings yet
- Spectrophotometric Determination of Unknown Bovine Serum Albumin (Alyza Joy M. Ramirez)Document5 pagesSpectrophotometric Determination of Unknown Bovine Serum Albumin (Alyza Joy M. Ramirez)Alyza Joy RamirezNo ratings yet
- Chapter 2 Leaf Structure and Function (Biology SPM)Document12 pagesChapter 2 Leaf Structure and Function (Biology SPM)ALAMEL MANZGHAI A/P GANESONNo ratings yet
- Synthego Ebook Crispr 101 PDFDocument29 pagesSynthego Ebook Crispr 101 PDFRie R100% (1)