You might also like
- AdisakwattanaS Etal 2012Document8 pagesAdisakwattanaS Etal 2012DhirajNo ratings yet
- Isolation and identification of terpenoids from chicory roots and their inhibitory activities against yeast α-glucosidaseDocument12 pagesIsolation and identification of terpenoids from chicory roots and their inhibitory activities against yeast α-glucosidaserini dwi wahyuningsihNo ratings yet
- 5a5b PDFDocument6 pages5a5b PDFIndah AfrianiNo ratings yet
- Antidiabetic Effect of Potential Indian Medicinal Plants: A Target Specific In-Vitro StudyDocument14 pagesAntidiabetic Effect of Potential Indian Medicinal Plants: A Target Specific In-Vitro StudyRajendra YegnambhatlaNo ratings yet
- Research Article: Stamineus Extract On Human Cytochrome P450Document7 pagesResearch Article: Stamineus Extract On Human Cytochrome P450Herry NavtantoNo ratings yet
- Ethanol Extract of Mahkota Dewa (Phaleria Macrocarpa (Scheff,) Boerl.)Document4 pagesEthanol Extract of Mahkota Dewa (Phaleria Macrocarpa (Scheff,) Boerl.)RisyadTorresAlamsyahNo ratings yet
- Evaluation of clonal herbs for managing diabetes and hypertensionDocument12 pagesEvaluation of clonal herbs for managing diabetes and hypertensionAlberto HMNo ratings yet
- Food Chemistry study identifies maltase inhibitors from flower budsDocument7 pagesFood Chemistry study identifies maltase inhibitors from flower budsRayyhana ShopNo ratings yet
- Efect Antihiperglicemiant 2.35 - 2013Document7 pagesEfect Antihiperglicemiant 2.35 - 2013Ienascu IoanaNo ratings yet
- Antioxidant and Antidiabetic Potential of Curcuma: Longa and Its CompoundsDocument13 pagesAntioxidant and Antidiabetic Potential of Curcuma: Longa and Its CompoundsMelati FitriaNo ratings yet
- 1 PBDocument6 pages1 PBUncu EchiNo ratings yet
- Research Article Vernonia Amygdalina: A Comparative StudyDocument14 pagesResearch Article Vernonia Amygdalina: A Comparative StudyNgan HoangNo ratings yet
- Industrial Crops and ProductsDocument11 pagesIndustrial Crops and ProductsYAYON PAMULA MUKTINo ratings yet
- 95687-Article Text-247599-1-10-20131023 PDFDocument10 pages95687-Article Text-247599-1-10-20131023 PDFJohn JosephNo ratings yet
- Sabrina Pharmaceuticals July 2023Document17 pagesSabrina Pharmaceuticals July 2023Ahmad AliNo ratings yet
- Summaries of ArticlesDocument5 pagesSummaries of ArticlesPharmanic By Ruttaba FatimaNo ratings yet
- In Vivo Study On The Efficacy of Hypoglycemic Activity of Spirulina Plantesis inDocument7 pagesIn Vivo Study On The Efficacy of Hypoglycemic Activity of Spirulina Plantesis ingallondNo ratings yet
- Setiadi 2019 J. Phys. Conf. Ser. 1146 012009Document8 pagesSetiadi 2019 J. Phys. Conf. Ser. 1146 012009Nelli Nur Indah SariNo ratings yet
- Hancornia speciosa Potential Anti-Diabetic EffectsDocument6 pagesHancornia speciosa Potential Anti-Diabetic EffectscleyslaNo ratings yet
- Antioxidant Activity and Anti-Hyperglycemic Effect Of: Lagenaria Siceraria Fruit ExtractDocument6 pagesAntioxidant Activity and Anti-Hyperglycemic Effect Of: Lagenaria Siceraria Fruit ExtractEgahasaniaNo ratings yet
- OB17 Siti Aisyah PDFDocument7 pagesOB17 Siti Aisyah PDFFakhri NasharulNo ratings yet
- Antidiabetic Activity of Ruellia Tuberosa L Role oDocument9 pagesAntidiabetic Activity of Ruellia Tuberosa L Role orandy hidayatNo ratings yet
- Inggris - SaponinDocument6 pagesInggris - SaponinrifaniNo ratings yet
- PHENOLIC, FLAVONOID AND DPPH RADICAL SCAVENGING ACTIVITYDocument3 pagesPHENOLIC, FLAVONOID AND DPPH RADICAL SCAVENGING ACTIVITYHana NurvitaNo ratings yet
- Journal of Functional Foods: Zi-Cheng Zhang, Hong-Bin Wang, Qing Zhou, Ben Hu, Jia-Hao Wen, Jiu-Liang ZhangDocument10 pagesJournal of Functional Foods: Zi-Cheng Zhang, Hong-Bin Wang, Qing Zhou, Ben Hu, Jia-Hao Wen, Jiu-Liang ZhangajengNo ratings yet
- 2022 Anti Dia Be TecDocument15 pages2022 Anti Dia Be TecHilda SarahNo ratings yet
- Antidiabetic Component SeparationDocument5 pagesAntidiabetic Component SeparationZarin juthiNo ratings yet
- 17 RJPT 17 2 2024Document7 pages17 RJPT 17 2 2024Imalia dwiputriNo ratings yet
- Umesh BJPR2 K10Document6 pagesUmesh BJPR2 K10ibrahimaNo ratings yet
- Sir Muqeet ArticleDocument12 pagesSir Muqeet Articlehassan khanNo ratings yet
- Evaluation of Antioxidant Activity of Clove Syzygium AromaticumDocument8 pagesEvaluation of Antioxidant Activity of Clove Syzygium AromaticumTrina ViskhawatNo ratings yet
- Comparative Evaluation of Polysaccharides Isolated From As-Tragalus, Oyster Mushroom, and Yacon As Inhibitors ofDocument4 pagesComparative Evaluation of Polysaccharides Isolated From As-Tragalus, Oyster Mushroom, and Yacon As Inhibitors ofGREESTYNo ratings yet
- Viscozyme L Aided Flavonoid Extraction and Identification of Quercetin From Saururus Chinensis (Lour.) BaillDocument5 pagesViscozyme L Aided Flavonoid Extraction and Identification of Quercetin From Saururus Chinensis (Lour.) BaillnguyendongthucNo ratings yet
- J. Phys.: Conf. Ser. 1940 012058 Hypoglycemic effects of Enhydra fluctuans aerial extractDocument8 pagesJ. Phys.: Conf. Ser. 1940 012058 Hypoglycemic effects of Enhydra fluctuans aerial extractrina delfitaNo ratings yet
- Joseph, 2013Document8 pagesJoseph, 2013Clarissa FrançaNo ratings yet
- Research Article: The Extract of Herbal Medicines Activates AMP-Activated Protein Kinase in Diet-Induced Obese RatsDocument9 pagesResearch Article: The Extract of Herbal Medicines Activates AMP-Activated Protein Kinase in Diet-Induced Obese RatsJoshNo ratings yet
- FILE 20210529 153215 ViewFreeArticleDocument2 pagesFILE 20210529 153215 ViewFreeArticleNguyen Thi Thuy Duong B1606703No ratings yet
- Efficacy of Aqueous Leaf Extract of Vernonia Amygdalina On Plasma Lipoprotein and Oxidative Status in Diabetic Rat Models. H. U. NwanjoDocument4 pagesEfficacy of Aqueous Leaf Extract of Vernonia Amygdalina On Plasma Lipoprotein and Oxidative Status in Diabetic Rat Models. H. U. NwanjoNgan HoangNo ratings yet
- IJPRAS (Manuscript) With Author InformationDocument12 pagesIJPRAS (Manuscript) With Author Informationkharpeaishwarya83No ratings yet
- Effects of Aronia polyphenols on oxidative stress and apoptosis in obese individualsDocument13 pagesEffects of Aronia polyphenols on oxidative stress and apoptosis in obese individualsLeszek Jaszczak0% (1)
- Asam Arakhidonat STZDocument6 pagesAsam Arakhidonat STZAlmas TNo ratings yet
- (Fullmethod) IJPSR15 06 12 102Document5 pages(Fullmethod) IJPSR15 06 12 102Zarin juthiNo ratings yet
- HTM 2015 Conference on Herbal and Traditional MedicineDocument8 pagesHTM 2015 Conference on Herbal and Traditional Medicineden mNo ratings yet
- Antidiabetic and Antihypercholesterolemic Activities of Citrus Sinensis Peel: in Vivo StudyDocument4 pagesAntidiabetic and Antihypercholesterolemic Activities of Citrus Sinensis Peel: in Vivo Studyneny21No ratings yet
- thử nghiệm ở chuộtDocument8 pagesthử nghiệm ở chuộtThanh NgânNo ratings yet
- Anti-Cancer and Antioxidant Properties of Phenolics Isolated From Toona Sinensis A Juss Acetone Leaf ExtractDocument9 pagesAnti-Cancer and Antioxidant Properties of Phenolics Isolated From Toona Sinensis A Juss Acetone Leaf ExtractGrace Anastasia Ginting SinusingaNo ratings yet
- Salfarina RamliDocument9 pagesSalfarina RamliMellisa Laura MintoroNo ratings yet
- Jurnal Kromatografi KertasDocument7 pagesJurnal Kromatografi Kertasmega rahmah utariNo ratings yet
- Phytochemical Profile, Antioxidative and Anti-Inflammatory Potentials of Gynura Bicolor DCDocument6 pagesPhytochemical Profile, Antioxidative and Anti-Inflammatory Potentials of Gynura Bicolor DCVanThanh DoNo ratings yet
- KonstipasiDocument6 pagesKonstipasiDiana Fitri MuslimahNo ratings yet
- Kayu ManisDocument6 pagesKayu ManisEca HadiputriNo ratings yet
- 500 - 1000 Pages For Book of Abstracts - I. INTERNATIONAL CONGRESS On MEDICINAL and AROMATIC PLANTS: "NATURAL and HEALTHY LIFE"Document500 pages500 - 1000 Pages For Book of Abstracts - I. INTERNATIONAL CONGRESS On MEDICINAL and AROMATIC PLANTS: "NATURAL and HEALTHY LIFE"Hasan Ibrahim Kozan50% (2)
- 1-s2.0-S2772371223000451-mainDocument7 pages1-s2.0-S2772371223000451-mainAmos Babu JettiNo ratings yet
- Chemical Nature, Stability and Bioefficacies of Anthocyanins From Fruit Peel of Syzygium Cumini Skeels PDFDocument9 pagesChemical Nature, Stability and Bioefficacies of Anthocyanins From Fruit Peel of Syzygium Cumini Skeels PDFMinyty LeNo ratings yet
- Antioxidant and Antidiabetic Potential of Galing Stem Extract (Cayratia Trifolia Domin)Document4 pagesAntioxidant and Antidiabetic Potential of Galing Stem Extract (Cayratia Trifolia Domin)Fauziah SyamsuddinNo ratings yet
- Anti Diabetic Activity of Ethanol Extract of Colocasia Esculenta Leaves in Alloxan Induced Diabetic RatsDocument4 pagesAnti Diabetic Activity of Ethanol Extract of Colocasia Esculenta Leaves in Alloxan Induced Diabetic RatsyigalbyNo ratings yet
- ANTIOXIDANT AND ANTI-DIABETIC POTENTIALDocument5 pagesANTIOXIDANT AND ANTI-DIABETIC POTENTIALSri LakshmiNo ratings yet
- My Solanum NigrumDocument11 pagesMy Solanum NigrumNadeemNo ratings yet
- Food Chemistry: Xiaoqing Yuan, Xiaohong Gu, Jian TangDocument6 pagesFood Chemistry: Xiaoqing Yuan, Xiaohong Gu, Jian TangHân Lê Thị MỹNo ratings yet
- Fruits, Vegetables, and Herbs: Bioactive Foods in Health PromotionFrom EverandFruits, Vegetables, and Herbs: Bioactive Foods in Health PromotionRating: 4 out of 5 stars4/5 (1)
- Cytotoxic Activity of Combretum Quadrangulare Leaf Extracts On Hepg2 Cancer Cell LineDocument3 pagesCytotoxic Activity of Combretum Quadrangulare Leaf Extracts On Hepg2 Cancer Cell LineNguyễn Minh AnhNo ratings yet
- VocabDocument3 pagesVocabNguyễn Minh AnhNo ratings yet
- Demora Is Lima 2012Document65 pagesDemora Is Lima 2012Nguyễn Minh AnhNo ratings yet
- Writing - Direct Question Essays - WS - 0Document3 pagesWriting - Direct Question Essays - WS - 0Nguyễn Minh AnhNo ratings yet
- Asami 2003Document3 pagesAsami 2003Nguyễn Minh AnhNo ratings yet
- TheDocument2 pagesTheNguyễn Minh AnhNo ratings yet
- Ielts Writing Task 1Document1 pageIelts Writing Task 1Nguyễn Minh AnhNo ratings yet
- Asami 2003Document3 pagesAsami 2003Nguyễn Minh AnhNo ratings yet
- VocabDocument3 pagesVocabNguyễn Minh AnhNo ratings yet
- (Alfazi) Câu Hỏi Trắc Nghiệm Từ Đồng Nghĩa - Trái NghĩaDocument12 pages(Alfazi) Câu Hỏi Trắc Nghiệm Từ Đồng Nghĩa - Trái NghĩaHuu DuyNo ratings yet
- Co So Hoa Hoc Di Vong Tran Quoc Son PDFDocument744 pagesCo So Hoa Hoc Di Vong Tran Quoc Son PDFNguyễn Minh AnhNo ratings yet
- (Huyennguyen) - Từ Vựng Thần ThôngDocument203 pages(Huyennguyen) - Từ Vựng Thần ThôngXuan Thuy TranNo ratings yet
- SAT Hot Words for Mastering VocabularyDocument13 pagesSAT Hot Words for Mastering VocabularystantonstudentNo ratings yet
- 7 Pin Dien Hoa Va The Dien Cuc 3uegiDocument4 pages7 Pin Dien Hoa Va The Dien Cuc 3uegiNguyễn Minh AnhNo ratings yet
- Sat Vocab PDFDocument100 pagesSat Vocab PDFIntekhab AlamNo ratings yet
- WA AP Chemistry Acid-Base Review #3 KeyDocument2 pagesWA AP Chemistry Acid-Base Review #3 KeyNguyễn Minh AnhNo ratings yet
- Exam3 EquationsheetDocument1 pageExam3 EquationsheetNguyễn Minh AnhNo ratings yet
- 0general Chemistry2-10Document32 pages0general Chemistry2-10Nguyễn Minh AnhNo ratings yet
- Chapter9Solutions and Solution ReactionsDocument25 pagesChapter9Solutions and Solution ReactionsdhavaleshNo ratings yet
- Worksheet 11 KeyDocument8 pagesWorksheet 11 KeyNguyễn Minh AnhNo ratings yet
- Practice Exam AnswersDocument19 pagesPractice Exam AnswersNguyễn Minh AnhNo ratings yet
- 1520 Solution 20 EquilibriaDocument25 pages1520 Solution 20 EquilibriaNguyễn Minh AnhNo ratings yet
- 0general Chemistry2-10Document32 pages0general Chemistry2-10Nguyễn Minh AnhNo ratings yet
- ChemChapter11 Mid Term2014Document24 pagesChemChapter11 Mid Term2014MaxwellNo ratings yet
- Inventory Storage and Retrieval System PatentDocument15 pagesInventory Storage and Retrieval System PatentdevanasokaNo ratings yet
- List of household items for relocationDocument4 pagesList of household items for relocationMADDYNo ratings yet
- Building Primitive Traps & SnaresDocument101 pagesBuilding Primitive Traps & SnaresJoseph Madr90% (10)
- PediculosisDocument14 pagesPediculosisREYMARK HACOSTA100% (1)
- 1571-1635319494618-Unit 04 Leadership and ManagementDocument48 pages1571-1635319494618-Unit 04 Leadership and ManagementdevindiNo ratings yet
- Sia Mock+Test 1 Csat Updated CompressedDocument216 pagesSia Mock+Test 1 Csat Updated Compressedpooja bhatiNo ratings yet
- One - Pager - SOGEVAC SV 320 BDocument2 pagesOne - Pager - SOGEVAC SV 320 BEOLOS COMPRESSORS LTDNo ratings yet
- Casestudy3 Hbo MaDocument2 pagesCasestudy3 Hbo Ma132345usdfghjNo ratings yet
- Barcelona Smart City TourDocument44 pagesBarcelona Smart City TourPepe JeansNo ratings yet
- Omobonike 1Document13 pagesOmobonike 1ODHIAMBO DENNISNo ratings yet
- Environmental Threats Differentiated Reading Comprehension Ver 1Document20 pagesEnvironmental Threats Differentiated Reading Comprehension Ver 1Camila DiasNo ratings yet
- History and Development of the Foodservice IndustryDocument23 pagesHistory and Development of the Foodservice IndustryMaria Athenna MallariNo ratings yet
- GRP Product CatalogueDocument57 pagesGRP Product CatalogueMulyana alcNo ratings yet
- Victor Mejia ResumeDocument1 pageVictor Mejia Resumeapi-510300922No ratings yet
- Virtio-Fs - A Shared File System For Virtual MachinesDocument21 pagesVirtio-Fs - A Shared File System For Virtual MachinesLeseldelaterreNo ratings yet
- TNTCL Cost Data 2021 22Document95 pagesTNTCL Cost Data 2021 22Akd DeshmukhNo ratings yet
- Plastic Welding: We Know HowDocument125 pagesPlastic Welding: We Know Howprabal rayNo ratings yet
- ProjectDocument86 pagesProjectrajuNo ratings yet
- INVENTORY MANAGEMENT TechniquesDocument24 pagesINVENTORY MANAGEMENT TechniquesWessal100% (1)
- Evolution Packet FinalDocument24 pagesEvolution Packet FinalJoaquinNo ratings yet
- K230F Equipment ManualsDocument166 pagesK230F Equipment ManualsHui ChenNo ratings yet
- WEEK 8. - Gen Math-PortfolioDocument12 pagesWEEK 8. - Gen Math-PortfolioOreo ProductionsNo ratings yet
- Understanding 2006 PSAT-NMSQT ScoresDocument8 pagesUnderstanding 2006 PSAT-NMSQT ScorestacobeoNo ratings yet
- Presepsi Khalayak Terhadap Program Acara Televise Reality Show "Jika Aku Menjadi" Di Trans TVDocument128 pagesPresepsi Khalayak Terhadap Program Acara Televise Reality Show "Jika Aku Menjadi" Di Trans TVAngga DianNo ratings yet
- Serv7107 V05N01 TXT7Document32 pagesServ7107 V05N01 TXT7azry_alqadry100% (6)
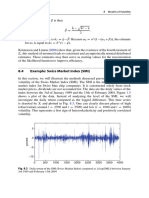
- 8.4 Example: Swiss Market Index (SMI) : 188 8 Models of VolatilityDocument3 pages8.4 Example: Swiss Market Index (SMI) : 188 8 Models of VolatilityNickesh ShahNo ratings yet
- Eagle Test ReportDocument25 pagesEagle Test ReportMuhammad FahadNo ratings yet
- How Do I Prepare For Public Administration For IAS by Myself Without Any Coaching? Which Books Should I Follow?Document3 pagesHow Do I Prepare For Public Administration For IAS by Myself Without Any Coaching? Which Books Should I Follow?saiviswanath0990100% (1)
- Service Positioning and DesignDocument3 pagesService Positioning and DesignSaurabh SinhaNo ratings yet
- General Biology 2-Week 2-Module 4-Evidence of EvolutionDocument16 pagesGeneral Biology 2-Week 2-Module 4-Evidence of EvolutionFEMALE Dawal LaizaNo ratings yet