You might also like
- 1 SMDocument29 pages1 SMrabacalabdoulNo ratings yet
- Bardet&Fernandez 2000 PDFDocument9 pagesBardet&Fernandez 2000 PDFEskizXNo ratings yet
- Theropod Paleobiology, More Than Just Bird Origins: Thomas R. HOLTZ, JRDocument3 pagesTheropod Paleobiology, More Than Just Bird Origins: Thomas R. HOLTZ, JRagomezgallardoNo ratings yet
- Antunes Mateus 2003 Dinosaurs of PortugalDocument19 pagesAntunes Mateus 2003 Dinosaurs of PortugalScribd_is_GreatNo ratings yet
- The Ancestry of Modern Amphibia: A Review of the EvidenceFrom EverandThe Ancestry of Modern Amphibia: A Review of the EvidenceNo ratings yet
- A Leopard Panthera Pardus L 1758 Find FRDocument11 pagesA Leopard Panthera Pardus L 1758 Find FRDanaGrujićNo ratings yet
- New Data On The Cainotheriidae (Mammalia, Artiodactyla) From The Early Oligocene of South-Western FranceDocument22 pagesNew Data On The Cainotheriidae (Mammalia, Artiodactyla) From The Early Oligocene of South-Western FranceCoster PAULINENo ratings yet
- Zoological Journal of The Linnean Society Volume 45 Issue 304 1964 (Doi 10.1111/j.1096-3642.1964.Tb00488.x) Robert L. Carroll - The Earliest ReptilesDocument24 pagesZoological Journal of The Linnean Society Volume 45 Issue 304 1964 (Doi 10.1111/j.1096-3642.1964.Tb00488.x) Robert L. Carroll - The Earliest ReptilesDiana MendoncaNo ratings yet
- The Mammals of Island EuropeDocument6 pagesThe Mammals of Island EuropeAca AvNo ratings yet
- Comments on the Taxonomic Status of Apodemus peninsulae, with Description of a New Subspecies from North ChinaFrom EverandComments on the Taxonomic Status of Apodemus peninsulae, with Description of a New Subspecies from North ChinaNo ratings yet
- A Maniraptoran Sacrum From The Upper Cretaceous of The HategDocument9 pagesA Maniraptoran Sacrum From The Upper Cretaceous of The HategJorge BarrientosNo ratings yet
- Beetles Insecta ColeopteraDocument14 pagesBeetles Insecta Coleopteradipeshsingh563No ratings yet
- The First Mandible of Gastornis Hébert, 1855 (Aves, Gastornithidae) From The Thanetian (Paleocene) of Mont-de-Berru (France)Document12 pagesThe First Mandible of Gastornis Hébert, 1855 (Aves, Gastornithidae) From The Thanetian (Paleocene) of Mont-de-Berru (France)Junior RubianoNo ratings yet
- The Ancestry of Modern Amphibia: A Review of the EvidenceFrom EverandThe Ancestry of Modern Amphibia: A Review of the EvidenceNo ratings yet
- Maastrichtian Dinosaurs in SW Transylvania (Romania), Codrea Et Al., 2010Document7 pagesMaastrichtian Dinosaurs in SW Transylvania (Romania), Codrea Et Al., 2010planessNo ratings yet
- Some Remarks On The African Wild AssDocument14 pagesSome Remarks On The African Wild AssFrancisco Javier Navas GonzálezNo ratings yet
- Dinosaurs Nesting On Tidal Flats: N. Lo Pez-Martı Nez A,, J.J. Moratalla B, J.L. Sanz BDocument11 pagesDinosaurs Nesting On Tidal Flats: N. Lo Pez-Martı Nez A,, J.J. Moratalla B, J.L. Sanz BManuel PérezNo ratings yet
- Trophon IaraeDocument4 pagesTrophon IaraeFede LópezNo ratings yet
- Gerold Steiner - The SnoutersDocument90 pagesGerold Steiner - The SnoutersMilan StefanNo ratings yet
- Paper BrachyopodaaDocument20 pagesPaper BrachyopodaaBimo KusumahastoNo ratings yet
- Jaguarundi Herpailurus Yagoaroundi Notes in The Caribbean Lowlands of Belize and Guatemala Mccarthy 1992Document5 pagesJaguarundi Herpailurus Yagoaroundi Notes in The Caribbean Lowlands of Belize and Guatemala Mccarthy 1992María José TorallaNo ratings yet
- The Ursus Spelaeus Disappearance ArchaeoDocument8 pagesThe Ursus Spelaeus Disappearance ArchaeoGianfranco BongioanniNo ratings yet
- Tubul FormationDocument16 pagesTubul FormationFernanda NarbonaNo ratings yet
- Pithecanthropus in Java: at at atDocument15 pagesPithecanthropus in Java: at at atAnth5334No ratings yet
- Mammuthus Primigenius in The Cave and Portable ArtDocument16 pagesMammuthus Primigenius in The Cave and Portable ArtMirela DragosNo ratings yet
- Geraads 2016 Pleistocene Carnivora (Mammalia) From Tighennif (Ternifine), AlgeriaDocument45 pagesGeraads 2016 Pleistocene Carnivora (Mammalia) From Tighennif (Ternifine), AlgeriaGhaier KazmiNo ratings yet
- Devonian Brachiopods of The Tamesna Basin (Central Sahara Algeria and North Niger) - Part 1Document52 pagesDevonian Brachiopods of The Tamesna Basin (Central Sahara Algeria and North Niger) - Part 1Carlos Alfredo Espinoza SullcaNo ratings yet
- Encyclopaedia Britannica, 11th Edition, Volume 8, Slice 10 "Echinoderma" to "Edward"From EverandEncyclopaedia Britannica, 11th Edition, Volume 8, Slice 10 "Echinoderma" to "Edward"No ratings yet
- Oldest Predaceous Water Bugs Insecta Heteroptera Belostomatidae With Implications For Paleolimnology of The Triassic Cow Branch FormationDocument12 pagesOldest Predaceous Water Bugs Insecta Heteroptera Belostomatidae With Implications For Paleolimnology of The Triassic Cow Branch FormationDinaNo ratings yet
- EISENMANN 1999 - Equus Granatensis Venta MicenaDocument15 pagesEISENMANN 1999 - Equus Granatensis Venta MicenaorcenseNo ratings yet
- Description of The Most Complete Skeleton of Stegomastodon (Mammalia, Gomphotheriidae) Recorded For The Mexican Late PleistoceneDocument18 pagesDescription of The Most Complete Skeleton of Stegomastodon (Mammalia, Gomphotheriidae) Recorded For The Mexican Late PleistoceneOctavio MoralesNo ratings yet
- The Relict Cerithiid Prosobranch, (Crosse) : Gourmya GourmyiDocument3 pagesThe Relict Cerithiid Prosobranch, (Crosse) : Gourmya GourmyiLukewarmNo ratings yet
- Pott, C. Et Al - Palaeontology - 2007Document20 pagesPott, C. Et Al - Palaeontology - 2007PaBloNo ratings yet
- 1997 Stephanorhinus Cf. Hundsheimensis (Toula) (Ridnocerotidae, Mammalia) From Trlica Near Pljevlja (Montenegro)Document15 pages1997 Stephanorhinus Cf. Hundsheimensis (Toula) (Ridnocerotidae, Mammalia) From Trlica Near Pljevlja (Montenegro)Predrag RadovićNo ratings yet
- Revision of The European Athecate Hydroids and Their Medusae (Hydrozoa, Cnidaria) : Families Oceanidae and PachycordylidaeDocument55 pagesRevision of The European Athecate Hydroids and Their Medusae (Hydrozoa, Cnidaria) : Families Oceanidae and PachycordylidaeRabha EnanehcNo ratings yet
- Of Archaeornithomimus Asiaticus OFDocument11 pagesOf Archaeornithomimus Asiaticus OFnomadNo ratings yet
- Donoghue 2001Document24 pagesDonoghue 2001J.D. NobleNo ratings yet
- The Pithecanthropus Erectus: Studies on Ancestral Stock of MankindFrom EverandThe Pithecanthropus Erectus: Studies on Ancestral Stock of MankindNo ratings yet
- History of The Dodo HumeDocument25 pagesHistory of The Dodo HumeEuro VillaNo ratings yet
- Description of Zabius Gaucho ScorpionesDocument11 pagesDescription of Zabius Gaucho ScorpionesGastón LópezNo ratings yet
- A Pictorial Atlas of Fossil Remains: Illustrations from Parkinson's "Organic remains of a former world," and Artis's "Antediluvian phytology"From EverandA Pictorial Atlas of Fossil Remains: Illustrations from Parkinson's "Organic remains of a former world," and Artis's "Antediluvian phytology"No ratings yet
- Angolatitan Adamastor, A New Sauropod Dinosaur: and The First Record From AngolaDocument13 pagesAngolatitan Adamastor, A New Sauropod Dinosaur: and The First Record From AngolaKanekoNo ratings yet
- Late Campanian Theropod Trackways From Porvenir de Jalpa, Coahuila, MexicoDocument7 pagesLate Campanian Theropod Trackways From Porvenir de Jalpa, Coahuila, MexicoNelson ValdesNo ratings yet
- Buthus BonitoDocument8 pagesButhus Bonitobibi boboNo ratings yet
- Four Remarkable From China: WenhuaDocument3 pagesFour Remarkable From China: WenhuaReticulatusNo ratings yet
- The Snouters - Form and Life of - Harald StumpkeDocument126 pagesThe Snouters - Form and Life of - Harald StumpkeAlberto Humanes MuñozNo ratings yet
- 1 s2.0 S1631068315002067 MainDocument12 pages1 s2.0 S1631068315002067 MainCarlos Saez MartinezNo ratings yet
- History of Humanity Hominid Rise CHDocument15 pagesHistory of Humanity Hominid Rise CHDurba GhoshNo ratings yet
- Bulanov & Sennikov 2006. The First Gliding Reptiles From The Upper Permian of RussiaDocument4 pagesBulanov & Sennikov 2006. The First Gliding Reptiles From The Upper Permian of RussiaИван БелоусовNo ratings yet
- Stenorhynchosaurus Munozi - VILLADELEYVADocument20 pagesStenorhynchosaurus Munozi - VILLADELEYVASERGIO MAYORGANo ratings yet
- First Record of Lingula (Brachiopoda) From The Miocene of France, With Diagnosis of L. DregeriDocument7 pagesFirst Record of Lingula (Brachiopoda) From The Miocene of France, With Diagnosis of L. DregeriJansius SaragihNo ratings yet
- The Freshwater Decapod Crustaceans of Halmahera, IndonesiaDocument32 pagesThe Freshwater Decapod Crustaceans of Halmahera, IndonesiaEdward WangNo ratings yet
- Csiki Et Al 2010 - PaludititanDocument28 pagesCsiki Et Al 2010 - PaludititanBruno NavarroNo ratings yet
- A Synopsis of Neotropical Hylid Frogs, Genus OsteocephalusFrom EverandA Synopsis of Neotropical Hylid Frogs, Genus OsteocephalusNo ratings yet
- Mammiferi Austria PleistoceneDocument13 pagesMammiferi Austria PleistoceneGianfranco BongioanniNo ratings yet
- LongDocument17 pagesLongChristine ElnasNo ratings yet
- List of Birds Collected in the Island of Bouru (One of the Moluccas), with Descriptions of the New SpeciesFrom EverandList of Birds Collected in the Island of Bouru (One of the Moluccas), with Descriptions of the New SpeciesNo ratings yet
- Pleistocene Pocket Gophers From San Josecito Cave, Nuevo Leon, MexicoFrom EverandPleistocene Pocket Gophers From San Josecito Cave, Nuevo Leon, MexicoNo ratings yet
- The Floating Drydock'S: Uss Alaska Cb1 ClassDocument69 pagesThe Floating Drydock'S: Uss Alaska Cb1 Classalex331994No ratings yet
- Portents of Protest in The Later Han Dynasty: The Memorials of Hsiang K'ai To Emperor Huan Rate de CrespignyDocument131 pagesPortents of Protest in The Later Han Dynasty: The Memorials of Hsiang K'ai To Emperor Huan Rate de Crespignyalex331994No ratings yet
- Recruitment Revisited For ANU Open ResearchDocument43 pagesRecruitment Revisited For ANU Open Researchalex331994No ratings yet
- Ra Va 3-04-20Document212 pagesRa Va 3-04-20alex331994No ratings yet
- I. Xun Yu and Xun You Amid Chaos: Cambridge History of China, Vol. 1, The Ch'in and Han Empires 221 BC - AD 220Document33 pagesI. Xun Yu and Xun You Amid Chaos: Cambridge History of China, Vol. 1, The Ch'in and Han Empires 221 BC - AD 220alex331994No ratings yet
- I. The Excellencies and AboveDocument20 pagesI. The Excellencies and Abovealex331994No ratings yet
- East Asian History: Papers On Far Eastern HistoryDocument27 pagesEast Asian History: Papers On Far Eastern Historyalex331994No ratings yet
- Gladius Brochure V7 PDFDocument12 pagesGladius Brochure V7 PDFalex331994No ratings yet
- Winged WolfDocument5 pagesWinged Wolfalex331994No ratings yet
- Player Guide 2.35 PDFDocument160 pagesPlayer Guide 2.35 PDFalex331994No ratings yet
- Brochure Eclipse PDFDocument7 pagesBrochure Eclipse PDFalex331994No ratings yet
- DCS F14 GuideDocument438 pagesDCS F14 Guidealex331994100% (4)
- Constellation Taurus V7Document12 pagesConstellation Taurus V7alex331994No ratings yet
- Freelancer Brochure PDFDocument20 pagesFreelancer Brochure PDFalex331994No ratings yet
- Redeemer Brochure V8Document12 pagesRedeemer Brochure V8alex331994100% (1)
- DUSTIN NUESTRO Skeletal System Lab. WorksheetDocument10 pagesDUSTIN NUESTRO Skeletal System Lab. WorksheetDustin NuestroNo ratings yet
- Nose AnatomyDocument113 pagesNose AnatomyPratiksha PatilNo ratings yet
- Biomechanics of Cervical SpineDocument25 pagesBiomechanics of Cervical SpineNithya NithyaNo ratings yet
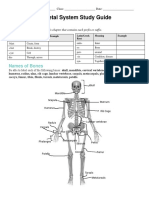
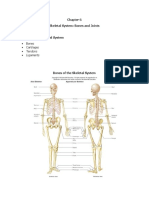
- Ethan McDaniel - Skeletal System Study GuideDocument7 pagesEthan McDaniel - Skeletal System Study GuideJohn BroNo ratings yet
- Laughing Is Also Good For Your Respiratory System!Document5 pagesLaughing Is Also Good For Your Respiratory System!A-Naeem To'mah Al-sawaieNo ratings yet
- 19 Norma Basalis DR GosaiDocument24 pages19 Norma Basalis DR GosaiDr.B.B.GosaiNo ratings yet
- Gross B Parotid Temporal and Infratemporal Regions Dr. JavierDocument6 pagesGross B Parotid Temporal and Infratemporal Regions Dr. JavierAngelo HuligangaNo ratings yet
- Pelvis 1Document10 pagesPelvis 1Nur Liyana MohamadNo ratings yet
- Skull Reviewer 2Document6 pagesSkull Reviewer 2Christian Bermas DioNo ratings yet
- Dimensi Tubuh Sapi Friesian Holstein Dan Limousin Betina Berdasarkan Morfometrik Dengan Citra DigitalDocument10 pagesDimensi Tubuh Sapi Friesian Holstein Dan Limousin Betina Berdasarkan Morfometrik Dengan Citra DigitalFawBlueNo ratings yet
- Posterior Subaxial Cervical Spine Screw Fixation: A Review of TechniquesDocument10 pagesPosterior Subaxial Cervical Spine Screw Fixation: A Review of TechniquesJaime XavierNo ratings yet
- Osteological ReproductionsDocument88 pagesOsteological ReproductionsAntonioNo ratings yet
- Bio FrogDocument4 pagesBio FrogAndrew John CellonaNo ratings yet
- VISCEROCRANIUMDocument157 pagesVISCEROCRANIUMTrần NamNo ratings yet
- Caldwell Luc - Radical Antrostomy - Procedure Canine Fossa and Inferior Meatal Puncture and Inferior Meatal AntrostomyDocument13 pagesCaldwell Luc - Radical Antrostomy - Procedure Canine Fossa and Inferior Meatal Puncture and Inferior Meatal AntrostomyAlexi Chávez EchevarríaNo ratings yet
- Axial Skeleton PDFDocument41 pagesAxial Skeleton PDFDewii FurryNo ratings yet
- HANDOUTS Prelim CH 6Document40 pagesHANDOUTS Prelim CH 6Abia Annieson A. LorenzoNo ratings yet
- Anatomy and Surgical Appraches of The Temporal Bone PDFDocument251 pagesAnatomy and Surgical Appraches of The Temporal Bone PDFThamescracker28809100% (2)
- Experiment 3Document3 pagesExperiment 3prathimapesaru7No ratings yet
- 2.osteology Q ADocument11 pages2.osteology Q ADr P N N ReddyNo ratings yet
- Labeled ItDocument50 pagesLabeled ItKaloy KamaoNo ratings yet
- VisibleBody MasticationDocument11 pagesVisibleBody MasticationdophuNo ratings yet
- 11 Axial Division of The Skeletal SystemDocument10 pages11 Axial Division of The Skeletal SystemMira Chariza Therese A. - BSN 1- CNo ratings yet
- Ks4 Bones Physical EducationDocument29 pagesKs4 Bones Physical EducationClaudia PinnaNo ratings yet
- Fibrous JointDocument5 pagesFibrous JointmamatnamakuNo ratings yet
- ENT Radiology ADocument64 pagesENT Radiology AMitulsinh M RavaljiNo ratings yet
- Chapter 04 Extraoral and Occlusal Anatomic LandmarksDocument22 pagesChapter 04 Extraoral and Occlusal Anatomic LandmarksFaith AnneessaNo ratings yet
- Advanced Surgical Facial Rejuvenation PDFDocument755 pagesAdvanced Surgical Facial Rejuvenation PDFFabiola Camberos Ruíz100% (1)
- Lab 4 - The Skeletal SystemDocument38 pagesLab 4 - The Skeletal SystemNatalie PembertonNo ratings yet
- 2 ArthrologyDocument30 pages2 ArthrologyAbhijith S. PNo ratings yet