You might also like
- A Simple Guide to Klippel-Feil Syndrome, Diagnosis, Treatment and Related ConditionsFrom EverandA Simple Guide to Klippel-Feil Syndrome, Diagnosis, Treatment and Related ConditionsNo ratings yet
- Panayiotopoulos Syndrome: A Common Benign But Underdiagnosed and Unexplored Early Childhood Seizure SyndromeDocument2 pagesPanayiotopoulos Syndrome: A Common Benign But Underdiagnosed and Unexplored Early Childhood Seizure SyndromeNensy AnggrainyNo ratings yet
- Benign Partial Epilepsy With Rolandic SpikesDocument4 pagesBenign Partial Epilepsy With Rolandic SpikesoxyNo ratings yet
- Angelman Syndrome - Identification and ManagementDocument10 pagesAngelman Syndrome - Identification and ManagementFajar YuniftiadiNo ratings yet
- 06 Neonatal SeizuresDocument10 pages06 Neonatal SeizuresSivanathan SubramaniamNo ratings yet
- Neonatal Seizures: Table 593-15 Measures in Clinical Practice To Reduce The Risk of SUDEPDocument13 pagesNeonatal Seizures: Table 593-15 Measures in Clinical Practice To Reduce The Risk of SUDEPmuhammad_ariefNo ratings yet
- AngelmanDocument12 pagesAngelmanapi-295680726No ratings yet
- Benign Epilepsies - Part 1Document35 pagesBenign Epilepsies - Part 1Azeem Ali ShahNo ratings yet
- E2. Epileptic SyndromesDocument15 pagesE2. Epileptic Syndromesyante88No ratings yet
- Mutations in Mitochondr Ial GenesDocument6 pagesMutations in Mitochondr Ial Geneslynden matbaganNo ratings yet
- Chapter 607 - Guillain-Barré Syndrome: Clinical ManifestationsDocument3 pagesChapter 607 - Guillain-Barré Syndrome: Clinical ManifestationsclaudiaNo ratings yet
- 5 Different Type of SyndromeDocument6 pages5 Different Type of SyndromeAdnan RezaNo ratings yet
- Seizure & Epilepsy PDFDocument11 pagesSeizure & Epilepsy PDFbencleese100% (1)
- Common Problems in Pediatric NeurologyDocument19 pagesCommon Problems in Pediatric NeurologyPaul ManaloNo ratings yet
- 586.1 Febrile Seizures Mohamad A. MikatiDocument5 pages586.1 Febrile Seizures Mohamad A. MikatiDewida 'dewet' MaulidatuNo ratings yet
- Neonatal SeizuresDocument11 pagesNeonatal SeizuresResmy Jose100% (1)
- Sharief F 2011Document14 pagesSharief F 2011wiwiNo ratings yet
- Classification: Epilepsy (Sometimes Referred To As A Seizure Disorder) Is A Common ChronicDocument11 pagesClassification: Epilepsy (Sometimes Referred To As A Seizure Disorder) Is A Common ChronicChristian BartolomeNo ratings yet
- Seziures in ChildhoodDocument45 pagesSeziures in ChildhoodLily AddamsNo ratings yet
- Bilateral Vestibulopathy BooKDocument6 pagesBilateral Vestibulopathy BooKmiranti fauziNo ratings yet
- NCBI Infantil Spasme PDFDocument5 pagesNCBI Infantil Spasme PDFRiri Dwi AnggaraniNo ratings yet
- Case 1Document2 pagesCase 1AyidhNo ratings yet
- Epilepsies Are Those Which Are Categorized by The Lobe in Which They Arise. Mesial Temporal Lobe Epilepsy (Or Hippocampal Epilepsy) May Be The MostDocument5 pagesEpilepsies Are Those Which Are Categorized by The Lobe in Which They Arise. Mesial Temporal Lobe Epilepsy (Or Hippocampal Epilepsy) May Be The MostvidushiiiNo ratings yet
- Complications MalariaDocument4 pagesComplications MalariaBenny Trisaktyari100% (1)
- Child NeurologyDocument8 pagesChild NeurologyANOOPVANo ratings yet
- Neonatalseizures:An Updateonmechanisms Andmanagement: Frances E. JensenDocument20 pagesNeonatalseizures:An Updateonmechanisms Andmanagement: Frances E. JensenJuan Martín Velasco CárdenasNo ratings yet
- Wernicke Korsakoff SyndromeDocument19 pagesWernicke Korsakoff Syndrometerry2wa100% (4)
- The Diagnosis and Management of Epilepsy: Journal of Tropical Pediatrics January 2002Document9 pagesThe Diagnosis and Management of Epilepsy: Journal of Tropical Pediatrics January 2002Roy MarechaNo ratings yet
- Infantile, Childhood, and Adolescent Epilepsies.9Document34 pagesInfantile, Childhood, and Adolescent Epilepsies.9Silviana IlieNo ratings yet
- Atypical Pres. of BCECTS-reviewDocument7 pagesAtypical Pres. of BCECTS-reviewAlexisNo ratings yet
- Dravet Syndrome and SCN1A Disorders - PaediatricFOAMDocument14 pagesDravet Syndrome and SCN1A Disorders - PaediatricFOAMshaik saifuddinNo ratings yet
- Epilepsy in ChildrenDocument34 pagesEpilepsy in ChildrenAdam MochtarNo ratings yet
- GB Syndrome and Other: Immune Mediated NeuropathiesDocument82 pagesGB Syndrome and Other: Immune Mediated NeuropathiesTeena ChandranNo ratings yet
- Fenilketonuria: Devi WuysangDocument23 pagesFenilketonuria: Devi WuysangChristantina Pradescha AssaNo ratings yet
- Frontal Lobe SeizuresDocument11 pagesFrontal Lobe Seizuresjhonny churata huarachiNo ratings yet
- Epilepsy: Incidence and PrevalenceDocument25 pagesEpilepsy: Incidence and Prevalenceshamma shahulhameedNo ratings yet
- Biology: Investigatory ProjectDocument22 pagesBiology: Investigatory ProjectNarayana BaipadithayaNo ratings yet
- Cerebellar DisordersDocument12 pagesCerebellar DisordersAnn SamNo ratings yet
- Hutchinson-Gilford Progeria Syndrome: ContentsDocument10 pagesHutchinson-Gilford Progeria Syndrome: ContentsVĩnh Phú PhạmNo ratings yet
- Cerebellar DisordersDocument16 pagesCerebellar DisordersNur Ajiyanti 'anthi' SabirinaNo ratings yet
- Investigatory Project inDocument31 pagesInvestigatory Project inrchandralega2205100% (1)
- Neonatal Epilepsy SyndromesDocument22 pagesNeonatal Epilepsy SyndromesJUAN CAMILO LANDAZURINo ratings yet
- Group 1 Seizure DisordersDocument13 pagesGroup 1 Seizure DisordersDiana MurguiaNo ratings yet
- Neonatal Seizures 2014Document17 pagesNeonatal Seizures 2014akankshaNo ratings yet
- Angelman Syndrome Is A Genetic Disorder That Primarily Affects The Nervous System. CharacteristicDocument4 pagesAngelman Syndrome Is A Genetic Disorder That Primarily Affects The Nervous System. CharacteristicFrancis Miko ManlangitNo ratings yet
- Down's Syndrome and Spina BifidaDocument23 pagesDown's Syndrome and Spina BifidaKajal KaliaNo ratings yet
- Gaucher 3 CDocument10 pagesGaucher 3 CSergio ZambranoNo ratings yet
- SturgeeWeber Syndrome From The Past To The Present YaDocument10 pagesSturgeeWeber Syndrome From The Past To The Present YaKathia AriasNo ratings yet
- Neonatal Seizures: Postgraduate Dept of PediatricsDocument42 pagesNeonatal Seizures: Postgraduate Dept of PediatricsG VenkateshNo ratings yet
- Dravet Syndrome FactsheetDocument1 pageDravet Syndrome FactsheetNational Coalition for Health Professional Education in GeneticsNo ratings yet
- Epilepsy and Developmental Disorders - Next Generation Sequencing in The ClinicDocument9 pagesEpilepsy and Developmental Disorders - Next Generation Sequencing in The ClinicLocklinaNo ratings yet
- Orphanet Journal of Rare Diseases: Jacobsen SyndromeDocument10 pagesOrphanet Journal of Rare Diseases: Jacobsen SyndromeMuhammad RezaNo ratings yet
- Re Ex Myoclonic Epilepsy of Infancy: Seizures Induced by Tactile StimulationDocument8 pagesRe Ex Myoclonic Epilepsy of Infancy: Seizures Induced by Tactile StimulationOziNo ratings yet
- Epilepsy and Loss of ConsciousnessDocument11 pagesEpilepsy and Loss of ConsciousnessMarwan M.No ratings yet
- Kejang Pada AnakDocument30 pagesKejang Pada AnakAmaliahHarumiKarimNo ratings yet
- The Benign Occipital Epilepsies of Childhood: An Overview of The Idiopathic Syndromes and of The Relationship To MigraineDocument15 pagesThe Benign Occipital Epilepsies of Childhood: An Overview of The Idiopathic Syndromes and of The Relationship To MigraineAjeng RiaNo ratings yet
- Genetic Disease in HumansDocument5 pagesGenetic Disease in Humanslastjoe71No ratings yet
- NeuroDocument123 pagesNeuroarchana singhNo ratings yet
- Wernicke's Encephalopathy - Lancet 2007-1Document14 pagesWernicke's Encephalopathy - Lancet 2007-1Fábio VinholyNo ratings yet
- Tuli Kongenital PDFDocument4 pagesTuli Kongenital PDFfauziahandnNo ratings yet
- Down Syndrome - Genetics PDFDocument6 pagesDown Syndrome - Genetics PDFHartotok VipnetNo ratings yet
- Down's Syndrome: Protection Against Cancer and The Therapeutic Potential of DSCR1Document4 pagesDown's Syndrome: Protection Against Cancer and The Therapeutic Potential of DSCR1baba ababNo ratings yet
- What People With Down Syndrome Can Teach Us About Cardiopulmonary DiseaseDocument16 pagesWhat People With Down Syndrome Can Teach Us About Cardiopulmonary DiseaseKishan ArrumugamNo ratings yet
- Angiogenic Profile Identifies Pulmonary Hypertension in Children With Down SyndromeDocument8 pagesAngiogenic Profile Identifies Pulmonary Hypertension in Children With Down Syndromebaba ababNo ratings yet
- Afp20141215p851 PDFDocument8 pagesAfp20141215p851 PDFViviane BarbosaNo ratings yet
- Down's Syndrome: Protection Against Cancer and The Therapeutic Potential of DSCR1Document4 pagesDown's Syndrome: Protection Against Cancer and The Therapeutic Potential of DSCR1baba ababNo ratings yet
- Angelman FRenPro90Document5 pagesAngelman FRenPro90AldriaBrandstoreNo ratings yet
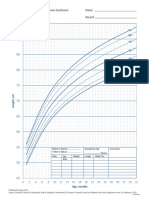
- DS Boys Length Birthto36mo PDFDocument1 pageDS Boys Length Birthto36mo PDFnisaNo ratings yet
- HHS Public Access: Endocrine Manifestations of Down SyndromeDocument11 pagesHHS Public Access: Endocrine Manifestations of Down Syndromebaba ababNo ratings yet
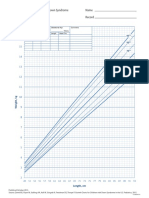
- DS Boys WeightLength Birthto36moDocument1 pageDS Boys WeightLength Birthto36moResti SyafitriNo ratings yet
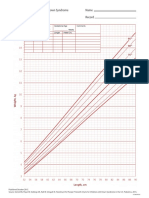
- DS Boys HeadCirc Birthto36moDocument1 pageDS Boys HeadCirc Birthto36moPeregrine Albertus Ricco AzaliNo ratings yet
- Neurocognition of Developmental Disorders of Language Ullman2017Document29 pagesNeurocognition of Developmental Disorders of Language Ullman2017baba ababNo ratings yet
- DS Girls WeightforLength Birthto36moDocument1 pageDS Girls WeightforLength Birthto36mobaba ababNo ratings yet
- Philpot Angelman Syn Cannabidiol RatDocument7 pagesPhilpot Angelman Syn Cannabidiol Ratbaba ababNo ratings yet
- Phonological Working Memory in Stuttering Bowers2018Document89 pagesPhonological Working Memory in Stuttering Bowers2018baba ababNo ratings yet
- Epilepsy in Angelman Syndrome: A Scoping Review: Debopam SamantaDocument13 pagesEpilepsy in Angelman Syndrome: A Scoping Review: Debopam Samantababa ababNo ratings yet
- Seizures and Angelman Syndrome Thiele Study ResultsDocument11 pagesSeizures and Angelman Syndrome Thiele Study Resultsbaba ababNo ratings yet
- Role of Executive Function in Developmental Stuttering Nihms-1062016Document19 pagesRole of Executive Function in Developmental Stuttering Nihms-1062016baba ababNo ratings yet
- Phonological Working Memory in Stuttering Bowers2018Document89 pagesPhonological Working Memory in Stuttering Bowers2018baba ababNo ratings yet
- COVID 19 Associated Vasculitis and Vasculopathy: Richard C. BeckerDocument13 pagesCOVID 19 Associated Vasculitis and Vasculopathy: Richard C. Beckerbaba ababNo ratings yet
- Normal Pressure Hydrocephalus Motor Apraxia Stutering Mathew2019Document17 pagesNormal Pressure Hydrocephalus Motor Apraxia Stutering Mathew2019baba ababNo ratings yet
- Neuromagnetic Oscillations in Sensory Systems Stuttering Epilepsy STB 2019Document13 pagesNeuromagnetic Oscillations in Sensory Systems Stuttering Epilepsy STB 2019baba ababNo ratings yet
- Stuttering Treatment and Its PharmacologyDocument8 pagesStuttering Treatment and Its Pharmacologybaba ababNo ratings yet
- Coagulop Endoteliop y Vasc CovidDocument9 pagesCoagulop Endoteliop y Vasc CovidbacchiocchipaoloNo ratings yet
- Kétnyelvőek Dadogása Hivatkozások Kornisch2021Document12 pagesKétnyelvőek Dadogása Hivatkozások Kornisch2021baba ababNo ratings yet
- Functional & Anatomical Bases of Developmental Stuttering 2019 Dec Nihms-1024485Document29 pagesFunctional & Anatomical Bases of Developmental Stuttering 2019 Dec Nihms-1024485baba ababNo ratings yet
- Developmental Stuttering Suppl Motor Cortex Busan2020Document15 pagesDevelopmental Stuttering Suppl Motor Cortex Busan2020baba ababNo ratings yet
- Cortico-Basal GGL Thalamcort Loop Development StutteringDocument15 pagesCortico-Basal GGL Thalamcort Loop Development Stutteringbaba ababNo ratings yet
- Cortico-Basal GGL Thalamcort Loop Development StutteringDocument15 pagesCortico-Basal GGL Thalamcort Loop Development Stutteringbaba ababNo ratings yet
- Lecture - Dr. Subagia - Neuro RadiologyDocument94 pagesLecture - Dr. Subagia - Neuro RadiologySyaimee Annisa AzzahraNo ratings yet
- HXX N3ur0anat0myyDocument29 pagesHXX N3ur0anat0myyLINDSLEY GONo ratings yet
- ChoreaDocument6 pagesChoreamainehoonaNo ratings yet
- Rett SyndromeDocument3 pagesRett Syndromeapi-240255006No ratings yet
- Gait and StationDocument17 pagesGait and Stationsarguss14100% (3)
- Hip Surveillance: Care PathwaysDocument2 pagesHip Surveillance: Care PathwaysMiguelNo ratings yet
- The Decompressive CraniectomyDocument32 pagesThe Decompressive CraniectomyAmerican English CourseNo ratings yet
- 2023 Aneurysmal Subarachnoid Hemorrhage Clinical Update SlidesDocument40 pages2023 Aneurysmal Subarachnoid Hemorrhage Clinical Update Slidesاحمد الشامي الشاميNo ratings yet
- Are Shaky Hands Normal or A Sign of Something - Piedmont HealthcareDocument4 pagesAre Shaky Hands Normal or A Sign of Something - Piedmont HealthcareLazarus BlackwellNo ratings yet
- Taiyang (M-Hn-9) : Sun (Supreme Yang)Document1 pageTaiyang (M-Hn-9) : Sun (Supreme Yang)ray72roNo ratings yet
- MKB Duffy Çeviri Ödevi-1.grupDocument62 pagesMKB Duffy Çeviri Ödevi-1.grupzhrksc1124No ratings yet
- Cerebral PalsyDocument9 pagesCerebral PalsyAngy100% (2)
- The Classification of Cerebral Palsy: Developmental Medicine & Child Neurology January 2005Document45 pagesThe Classification of Cerebral Palsy: Developmental Medicine & Child Neurology January 2005SuryanrNo ratings yet
- Logsheet 2018Document66 pagesLogsheet 2018Setiawati PinemNo ratings yet
- Conus Medullaris Vs Cauda Equina SyndromeDocument1 pageConus Medullaris Vs Cauda Equina SyndromecpradheepNo ratings yet
- AdrenoleukodystrophyDocument10 pagesAdrenoleukodystrophyapi-485321550No ratings yet
- Nicolet VikingQuest PageDocument2 pagesNicolet VikingQuest PageoviNo ratings yet
- 4 - Management of Patients With Neurologic Dysfunction Origenal K.Document21 pages4 - Management of Patients With Neurologic Dysfunction Origenal K.Taif SalimNo ratings yet
- Cerebral HemorrhageDocument10 pagesCerebral HemorrhageJayd Lorenz Vicente ChuanNo ratings yet
- Tumours of The Central Nervous System: FM Brett MD., FrcpathDocument57 pagesTumours of The Central Nervous System: FM Brett MD., FrcpathRerendhutNo ratings yet
- 10 2 3 29 EDSS - FormDocument2 pages10 2 3 29 EDSS - FormChibulcuteanNo ratings yet
- Impaired Verbal CommunicationDocument3 pagesImpaired Verbal CommunicationDesiree Deleon Guerrero0% (2)
- Neuromodulation NotesDocument7 pagesNeuromodulation NotesShashankChaurasiaNo ratings yet
- Pathological GaitDocument12 pagesPathological GaitNwa NnaNo ratings yet
- Frankel ScoreDocument5 pagesFrankel ScoreaaaNo ratings yet
- Comorbidities and Their RelatiDocument5 pagesComorbidities and Their RelatiDwi juliana DewiNo ratings yet
- Neurofibromatosis - Pediatrics - MSD Manual Professional EditionDocument8 pagesNeurofibromatosis - Pediatrics - MSD Manual Professional Editioncynthiadamayanti_drNo ratings yet
- Struktur Organisasi Neurologi 18 Mei 2020Document36 pagesStruktur Organisasi Neurologi 18 Mei 2020MienaNo ratings yet
- GSAQDocument1 pageGSAQGiga HasabiNo ratings yet
- Ictal and Postictal Psychiatric Disturbances - TrimbleDocument5 pagesIctal and Postictal Psychiatric Disturbances - Trimblegoo99No ratings yet
- An Autobiography of Trauma: A Healing JourneyFrom EverandAn Autobiography of Trauma: A Healing JourneyRating: 5 out of 5 stars5/5 (2)
- Summary: The Myth of Normal: Trauma, Illness, and Healing in a Toxic Culture By Gabor Maté MD & Daniel Maté: Key Takeaways, Summary & AnalysisFrom EverandSummary: The Myth of Normal: Trauma, Illness, and Healing in a Toxic Culture By Gabor Maté MD & Daniel Maté: Key Takeaways, Summary & AnalysisRating: 4 out of 5 stars4/5 (9)
- The Complex PTSD Workbook: A Mind-Body Approach to Regaining Emotional Control & Becoming WholeFrom EverandThe Complex PTSD Workbook: A Mind-Body Approach to Regaining Emotional Control & Becoming WholeRating: 4.5 out of 5 stars4.5/5 (49)
- My Grandmother's Hands: Racialized Trauma and the Pathway to Mending Our Hearts and BodiesFrom EverandMy Grandmother's Hands: Racialized Trauma and the Pathway to Mending Our Hearts and BodiesRating: 5 out of 5 stars5/5 (70)
- Rewire Your Anxious Brain: How to Use the Neuroscience of Fear to End Anxiety, Panic, and WorryFrom EverandRewire Your Anxious Brain: How to Use the Neuroscience of Fear to End Anxiety, Panic, and WorryRating: 4.5 out of 5 stars4.5/5 (157)
- The Upward Spiral: Using Neuroscience to Reverse the Course of Depression, One Small Change at a TimeFrom EverandThe Upward Spiral: Using Neuroscience to Reverse the Course of Depression, One Small Change at a TimeRating: 4.5 out of 5 stars4.5/5 (140)
- Summary of The Body Keeps the Score: Brain, Mind, and Body in the Healing of Trauma by Bessel van der Kolk MDFrom EverandSummary of The Body Keeps the Score: Brain, Mind, and Body in the Healing of Trauma by Bessel van der Kolk MDRating: 4.5 out of 5 stars4.5/5 (167)
- Vagus Nerve: A Complete Self Help Guide to Stimulate and Activate Vagal Tone — A Self Healing Exercises to Reduce Chronic Illness, PTSD, Anxiety, Inflammation, Depression, Trauma, and AngerFrom EverandVagus Nerve: A Complete Self Help Guide to Stimulate and Activate Vagal Tone — A Self Healing Exercises to Reduce Chronic Illness, PTSD, Anxiety, Inflammation, Depression, Trauma, and AngerRating: 4.5 out of 5 stars4.5/5 (16)
- Rapid Weight Loss Hypnosis: How to Lose Weight with Self-Hypnosis, Positive Affirmations, Guided Meditations, and Hypnotherapy to Stop Emotional Eating, Food Addiction, Binge Eating and MoreFrom EverandRapid Weight Loss Hypnosis: How to Lose Weight with Self-Hypnosis, Positive Affirmations, Guided Meditations, and Hypnotherapy to Stop Emotional Eating, Food Addiction, Binge Eating and MoreRating: 5 out of 5 stars5/5 (17)
- The Worry Trick: How Your Brain Tricks You into Expecting the Worst and What You Can Do About ItFrom EverandThe Worry Trick: How Your Brain Tricks You into Expecting the Worst and What You Can Do About ItRating: 4.5 out of 5 stars4.5/5 (107)
- Somatic Therapy Workbook: A Step-by-Step Guide to Experiencing Greater Mind-Body ConnectionFrom EverandSomatic Therapy Workbook: A Step-by-Step Guide to Experiencing Greater Mind-Body ConnectionNo ratings yet
- Don't Panic: Taking Control of Anxiety AttacksFrom EverandDon't Panic: Taking Control of Anxiety AttacksRating: 4 out of 5 stars4/5 (12)
- BORDERLINE PERSONALITY DISORDER: Help Yourself and Help Others. Articulate Guide to BPD. Tools and Techniques to Control Emotions, Anger, and Mood Swings. Save All Your Relationships and Yourself. NEW VERSIONFrom EverandBORDERLINE PERSONALITY DISORDER: Help Yourself and Help Others. Articulate Guide to BPD. Tools and Techniques to Control Emotions, Anger, and Mood Swings. Save All Your Relationships and Yourself. NEW VERSIONRating: 4.5 out of 5 stars4.5/5 (24)
- Brain Inflamed: Uncovering the Hidden Causes of Anxiety, Depression, and Other Mood Disorders in Adolescents and TeensFrom EverandBrain Inflamed: Uncovering the Hidden Causes of Anxiety, Depression, and Other Mood Disorders in Adolescents and TeensRating: 5 out of 5 stars5/5 (2)
- Summary: No Bad Parts: Healing Trauma and Restoring Wholeness with the Internal Family Systems Model by Richard C. Schwartz PhD & Alanis Morissette: Key Takeaways, Summary & AnalysisFrom EverandSummary: No Bad Parts: Healing Trauma and Restoring Wholeness with the Internal Family Systems Model by Richard C. Schwartz PhD & Alanis Morissette: Key Takeaways, Summary & AnalysisRating: 5 out of 5 stars5/5 (5)
- Binaural Beats: Activation of pineal gland – Stress reduction – Meditation – Brainwave entrainment – Deep relaxationFrom EverandBinaural Beats: Activation of pineal gland – Stress reduction – Meditation – Brainwave entrainment – Deep relaxationRating: 5 out of 5 stars5/5 (9)
- A Profession Without Reason: The Crisis of Contemporary Psychiatry—Untangled and Solved by Spinoza, Freethinking, and Radical EnlightenmentFrom EverandA Profession Without Reason: The Crisis of Contemporary Psychiatry—Untangled and Solved by Spinoza, Freethinking, and Radical EnlightenmentNo ratings yet
- Critical Thinking: How to Effectively Reason, Understand Irrationality, and Make Better DecisionsFrom EverandCritical Thinking: How to Effectively Reason, Understand Irrationality, and Make Better DecisionsRating: 4.5 out of 5 stars4.5/5 (39)
- Heal the Body, Heal the Mind: A Somatic Approach to Moving Beyond TraumaFrom EverandHeal the Body, Heal the Mind: A Somatic Approach to Moving Beyond TraumaRating: 4.5 out of 5 stars4.5/5 (56)
- Winning the War in Your Mind: Change Your Thinking, Change Your LifeFrom EverandWinning the War in Your Mind: Change Your Thinking, Change Your LifeRating: 5 out of 5 stars5/5 (558)
- Redefining Anxiety: What It Is, What It Isn't, and How to Get Your Life BackFrom EverandRedefining Anxiety: What It Is, What It Isn't, and How to Get Your Life BackRating: 4.5 out of 5 stars4.5/5 (152)
- When Religion Hurts You: Healing from Religious Trauma and the Impact of High-Control ReligionFrom EverandWhen Religion Hurts You: Healing from Religious Trauma and the Impact of High-Control ReligionRating: 4 out of 5 stars4/5 (11)
- I Don't Want to Talk About It: Overcoming the Secret Legacy of Male DepressionFrom EverandI Don't Want to Talk About It: Overcoming the Secret Legacy of Male DepressionRating: 4.5 out of 5 stars4.5/5 (106)
- Feel the Fear… and Do It Anyway: Dynamic Techniques for Turning Fear, Indecision, and Anger into Power, Action, and LoveFrom EverandFeel the Fear… and Do It Anyway: Dynamic Techniques for Turning Fear, Indecision, and Anger into Power, Action, and LoveRating: 4 out of 5 stars4/5 (250)