You might also like
- Cutaneous Silent Periods - Part 1 - Update On Physiological MechanismsDocument16 pagesCutaneous Silent Periods - Part 1 - Update On Physiological MechanismsGogy MarkNo ratings yet
- Progress in Neurobiology: Joana Cabral, Morten L. Kringelbach, Gustavo DecoDocument30 pagesProgress in Neurobiology: Joana Cabral, Morten L. Kringelbach, Gustavo DecoJavierElectricoNo ratings yet
- The Thalamocortical Vestibular System in Animals and HumansDocument28 pagesThe Thalamocortical Vestibular System in Animals and HumansCabinet VeterinarNo ratings yet
- Ain Stimulation in The Study and Treatment of AddictionDocument16 pagesAin Stimulation in The Study and Treatment of Addictionmariobar17636No ratings yet
- Waiting to Perceive: The Neurophysiology of Reward and Punishment AnticipationDocument11 pagesWaiting to Perceive: The Neurophysiology of Reward and Punishment AnticipationsoylahijadeunvampiroNo ratings yet
- Yawning-Its Anatomy, Chemistry, Role, and Pathological Considerations 2018Document18 pagesYawning-Its Anatomy, Chemistry, Role, and Pathological Considerations 2018Gilbert ThierryNo ratings yet
- Cutaneous Silent Periods - Part 2 - Update On Pathophysiology and Clinical UtilityDocument12 pagesCutaneous Silent Periods - Part 2 - Update On Pathophysiology and Clinical UtilityGogy MarkNo ratings yet
- Duffau (2018) - Error de BrocaDocument9 pagesDuffau (2018) - Error de BrocaMaría Cordero Varela0% (1)
- 1 s2.0 S0166432811007492 MainDocument5 pages1 s2.0 S0166432811007492 MainneuropsicojcNo ratings yet
- Progress in Neurobiology: G. Taccola, D. Sayenko, P. Gad, Y. Gerasimenko, V.R. EdgertonDocument18 pagesProgress in Neurobiology: G. Taccola, D. Sayenko, P. Gad, Y. Gerasimenko, V.R. EdgertonLeonardo CordeiroNo ratings yet
- Behavioural and Neurochemical Effects of Post-Weaning Social Isolation inDocument16 pagesBehavioural and Neurochemical Effects of Post-Weaning Social Isolation inNityananda PortelladaNo ratings yet
- Emotion and Motivation: The Role of The Amygdala, Ventral Striatum, and Prefrontal CortexDocument32 pagesEmotion and Motivation: The Role of The Amygdala, Ventral Striatum, and Prefrontal Cortexaragorn26No ratings yet
- Insulin-Mediated Synaptic Plasticity in The CNSDocument10 pagesInsulin-Mediated Synaptic Plasticity in The CNSAldehydeNo ratings yet
- KPCPDocument148 pagesKPCPstay312No ratings yet
- Functions Offlavonoids in the Central Nervous System - Astrocytes Astargets for Natural Compounds - IMAGEM FLAVONOIDESDocument7 pagesFunctions Offlavonoids in the Central Nervous System - Astrocytes Astargets for Natural Compounds - IMAGEM FLAVONOIDESrosybioNo ratings yet
- The Neuropsychology of Basal Ganglia: Review ArticleDocument6 pagesThe Neuropsychology of Basal Ganglia: Review ArticleLuisa FernandaNo ratings yet
- (2019) Kiernan MC. Measurement of Axonal Excitability - Consensus GuidelinesDocument16 pages(2019) Kiernan MC. Measurement of Axonal Excitability - Consensus GuidelinesLucía del ValleNo ratings yet
- TMP 346 CDocument10 pagesTMP 346 CFrontiersNo ratings yet
- Strategies For Repair and Regeneration: Eural Issue NgineeringDocument62 pagesStrategies For Repair and Regeneration: Eural Issue NgineeringVicente JordanNo ratings yet
- Cardiac, OptogeneticsDocument11 pagesCardiac, OptogeneticshisenknNo ratings yet
- 2015 An in Depth View of Avian SleepDocument8 pages2015 An in Depth View of Avian SleeppsicosmosNo ratings yet
- Iulia M Comsa - Tracking Brain Dynamics Across Transitions of ConsciousnessDocument150 pagesIulia M Comsa - Tracking Brain Dynamics Across Transitions of ConsciousnessIulia M ComsaNo ratings yet
- Neural Reuse of Action Perception Circuits For Language, Concepts and CommunicationDocument44 pagesNeural Reuse of Action Perception Circuits For Language, Concepts and CommunicationraamonNo ratings yet
- Ax Mac Her 2006 MemoryDocument13 pagesAx Mac Her 2006 MemorypastafarianboyNo ratings yet
- Blood Brain BarrierDocument5 pagesBlood Brain BarrierDaniel Felipe SantosNo ratings yet
- Neural Mechanism of AcupunctureDocument21 pagesNeural Mechanism of Acupunctureoh_leninhaNo ratings yet
- Emotional Tagging A Simple Hypothesis inDocument13 pagesEmotional Tagging A Simple Hypothesis inpyong jNo ratings yet
- John, E.R. (2002) - The Neurophysics of Consciousness. Brain Research Reviews 39, 1-28Document28 pagesJohn, E.R. (2002) - The Neurophysics of Consciousness. Brain Research Reviews 39, 1-28PaulNo ratings yet
- Astrocytes Multitaskers in Chronic PainDocument9 pagesAstrocytes Multitaskers in Chronic PainPedro FonsecaNo ratings yet
- Qasim Columbia 0054D 16299Document152 pagesQasim Columbia 0054D 16299Francisco OlivaresNo ratings yet
- Central Mechanisms in Phantom Limb PerceptionDocument14 pagesCentral Mechanisms in Phantom Limb PerceptionKarin Valenzuela100% (1)
- Neuroscience of Improvisation 2015Document10 pagesNeuroscience of Improvisation 2015Cassia LmtNo ratings yet
- Bhalsing 2013Document5 pagesBhalsing 2013Pradnya ParamitaNo ratings yet
- Neuroimage: ReviewDocument12 pagesNeuroimage: ReviewxgdfgmNo ratings yet
- Role of The Arcuate Nucleus of The Hypothalamus in Regulation of Body Weightduring Energy DeficitDocument11 pagesRole of The Arcuate Nucleus of The Hypothalamus in Regulation of Body Weightduring Energy DeficitJaime GonzálezNo ratings yet
- Descending ControlDocument120 pagesDescending ControlJessica Alejandra Pizarro MoralesNo ratings yet
- Ejercicio Cognición y NeuroplasticidadDocument15 pagesEjercicio Cognición y NeuroplasticidadcornytyNo ratings yet
- 1 s2.0 S2467981X17300100 MainDocument5 pages1 s2.0 S2467981X17300100 MainGian Franco Oneto TapiaNo ratings yet
- Developmental Cognitive Neuroscience: Mark H. JohnsonDocument7 pagesDevelopmental Cognitive Neuroscience: Mark H. JohnsonoconbhuiNo ratings yet
- Articulo Variaciones Anatomicas Nervio MaxilarDocument12 pagesArticulo Variaciones Anatomicas Nervio MaxilarCamila MaceaNo ratings yet
- Nicoletti 2007Document172 pagesNicoletti 2007Vladimir ShokarNo ratings yet
- Baslet 2011Document13 pagesBaslet 2011esckere papas fritasNo ratings yet
- Silva 2014Document34 pagesSilva 2014Oiy Saiank StabilNo ratings yet
- Non Invasive Human Brain StimulationDocument41 pagesNon Invasive Human Brain StimulationnarkeeshNo ratings yet
- Hijacking the mind: The tricks and strategies used to control your mind and how to break freeFrom EverandHijacking the mind: The tricks and strategies used to control your mind and how to break freeNo ratings yet
- ProceedingsELAINE 2020Document64 pagesProceedingsELAINE 2020farooqi111No ratings yet
- Nombela 2013 Into The Groove Can Rhythm Influence Parkinson's Disease PDFDocument7 pagesNombela 2013 Into The Groove Can Rhythm Influence Parkinson's Disease PDFXincrosisLemuriaNo ratings yet
- Fuad Lechin, Bertha Van Der Dijs, Gerardo Herna Ndez-Adria NDocument21 pagesFuad Lechin, Bertha Van Der Dijs, Gerardo Herna Ndez-Adria NMaria Lucrecia CrespoNo ratings yet
- Neuroscience and Biobehavioral Reviews: Mortimer MamelakDocument17 pagesNeuroscience and Biobehavioral Reviews: Mortimer MamelakDaniela Cruz MartinezNo ratings yet
- J Jelekin 2010 08 009 PDFDocument12 pagesJ Jelekin 2010 08 009 PDFRommel AndresNo ratings yet
- tmp22FA TMPDocument9 pagestmp22FA TMPFrontiersNo ratings yet
- Grossberg, Stephen (2017) - Towards Solving The Hard Problem of Consciousness - The Varieties of Brain Resonances and The Conscious Experiences That They Support. Neural Networks, 87, 38-95.Document58 pagesGrossberg, Stephen (2017) - Towards Solving The Hard Problem of Consciousness - The Varieties of Brain Resonances and The Conscious Experiences That They Support. Neural Networks, 87, 38-95.mindqualiaNo ratings yet
- Ideomotor Apraxia: A Review: Lewis A. Wheaton, Mark HallettDocument10 pagesIdeomotor Apraxia: A Review: Lewis A. Wheaton, Mark HallettCamila MadridNo ratings yet
- PD treatment dyskinesia: Disease or drug featureDocument9 pagesPD treatment dyskinesia: Disease or drug featureArio AkbarNo ratings yet
- Noninvasive Human Brain Stimulation: Timothy Wagner, Antoni Valero-Cabre, and Alvaro Pascual-LeoneDocument41 pagesNoninvasive Human Brain Stimulation: Timothy Wagner, Antoni Valero-Cabre, and Alvaro Pascual-LeoneGhinea IulianaNo ratings yet
- Electrified Minds Transcranial Direct Current Stimulation (TDCS) and GalvanicDocument22 pagesElectrified Minds Transcranial Direct Current Stimulation (TDCS) and GalvanicDanish MujibNo ratings yet
- Physiology & Behavior: Ian C. Webb, Michael N. Lehman, Lique M. CoolenDocument12 pagesPhysiology & Behavior: Ian C. Webb, Michael N. Lehman, Lique M. Coolencarlos orjuelaNo ratings yet
- Repetitive Transcranial Magnetic Stimulation For Neuropsychiatric DisordersDocument7 pagesRepetitive Transcranial Magnetic Stimulation For Neuropsychiatric DisordersMuskaan KhannaNo ratings yet
- Regional Anesthesia in CattleDocument33 pagesRegional Anesthesia in CattleBibek SutradharNo ratings yet
- 1 s2.0 S1568997213000360 MainDocument12 pages1 s2.0 S1568997213000360 MainValiant CheungNo ratings yet
- Angiogenic profile identifies pulmonary hypertension in children with Down syndromeDocument8 pagesAngiogenic profile identifies pulmonary hypertension in children with Down syndromebaba ababNo ratings yet
- Down's Syndrome: Protection Against Cancer and The Therapeutic Potential of DSCR1Document4 pagesDown's Syndrome: Protection Against Cancer and The Therapeutic Potential of DSCR1baba ababNo ratings yet
- Down's Syndrome: Protection Against Cancer and The Therapeutic Potential of DSCR1Document4 pagesDown's Syndrome: Protection Against Cancer and The Therapeutic Potential of DSCR1baba ababNo ratings yet
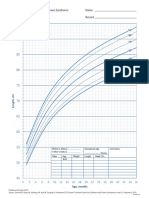
- DS Girls WeightforLength Birthto36moDocument1 pageDS Girls WeightforLength Birthto36mobaba ababNo ratings yet
- Afp20141215p851 PDFDocument8 pagesAfp20141215p851 PDFViviane BarbosaNo ratings yet
- DS Boys WeightLength Birthto36moDocument1 pageDS Boys WeightLength Birthto36moResti SyafitriNo ratings yet
- HHS Public Access: Endocrine Manifestations of Down SyndromeDocument11 pagesHHS Public Access: Endocrine Manifestations of Down Syndromebaba ababNo ratings yet
- What People With Down Syndrome Can Teach Us About Cardiopulmonary DiseaseDocument16 pagesWhat People With Down Syndrome Can Teach Us About Cardiopulmonary DiseaseKishan ArrumugamNo ratings yet
- Philpot Angelman Syn Cannabidiol RatDocument7 pagesPhilpot Angelman Syn Cannabidiol Ratbaba ababNo ratings yet
- Down Syndrome - Genetics PDFDocument6 pagesDown Syndrome - Genetics PDFHartotok VipnetNo ratings yet
- Angelman FRenPro90Document5 pagesAngelman FRenPro90AldriaBrandstoreNo ratings yet
- Phonological Working Memory in Stuttering Bowers2018Document89 pagesPhonological Working Memory in Stuttering Bowers2018baba ababNo ratings yet
- Seizures and Angelman Syndrome Thiele Study ResultsDocument11 pagesSeizures and Angelman Syndrome Thiele Study Resultsbaba ababNo ratings yet
- Growth Charts for Boys with Down Syndrome Birth to 36 MonthsDocument1 pageGrowth Charts for Boys with Down Syndrome Birth to 36 MonthsnisaNo ratings yet
- DS Boys HeadCirc Birthto36moDocument1 pageDS Boys HeadCirc Birthto36moPeregrine Albertus Ricco AzaliNo ratings yet
- Epilepsy in Angelman Syndrome: A Scoping Review: Debopam SamantaDocument13 pagesEpilepsy in Angelman Syndrome: A Scoping Review: Debopam Samantababa ababNo ratings yet
- Epilepsy in Angelman Syndrome: Karine Pelc, Stewart G. Boyd, Guy Cheron, Bernard DanDocument7 pagesEpilepsy in Angelman Syndrome: Karine Pelc, Stewart G. Boyd, Guy Cheron, Bernard Danbaba ababNo ratings yet
- Stuttering Treatment and Its PharmacologyDocument8 pagesStuttering Treatment and Its Pharmacologybaba ababNo ratings yet
- Phonological Working Memory in Stuttering Bowers2018Document89 pagesPhonological Working Memory in Stuttering Bowers2018baba ababNo ratings yet
- COVID 19 Associated Vasculitis and Vasculopathy: Richard C. BeckerDocument13 pagesCOVID 19 Associated Vasculitis and Vasculopathy: Richard C. Beckerbaba ababNo ratings yet
- Functional & Anatomical Bases of Developmental Stuttering 2019 Dec Nihms-1024485Document29 pagesFunctional & Anatomical Bases of Developmental Stuttering 2019 Dec Nihms-1024485baba ababNo ratings yet
- Normal Pressure Hydrocephalus Motor Apraxia Stutering Mathew2019Document17 pagesNormal Pressure Hydrocephalus Motor Apraxia Stutering Mathew2019baba ababNo ratings yet
- Role of Executive Function in Developmental Stuttering Nihms-1062016Document19 pagesRole of Executive Function in Developmental Stuttering Nihms-1062016baba ababNo ratings yet
- Coagulop Endoteliop y Vasc CovidDocument9 pagesCoagulop Endoteliop y Vasc CovidbacchiocchipaoloNo ratings yet
- Neurocognition of Developmental Disorders of Language Ullman2017Document29 pagesNeurocognition of Developmental Disorders of Language Ullman2017baba ababNo ratings yet
- Developmental Stuttering Suppl Motor Cortex Busan2020Document15 pagesDevelopmental Stuttering Suppl Motor Cortex Busan2020baba ababNo ratings yet
- Cortico-Basal GGL Thalamcort Loop Development StutteringDocument15 pagesCortico-Basal GGL Thalamcort Loop Development Stutteringbaba ababNo ratings yet
- Cortico-Basal GGL Thalamcort Loop Development StutteringDocument15 pagesCortico-Basal GGL Thalamcort Loop Development Stutteringbaba ababNo ratings yet
- Kétnyelvőek Dadogása Hivatkozások Kornisch2021Document12 pagesKétnyelvőek Dadogása Hivatkozások Kornisch2021baba ababNo ratings yet
- FEARS Database-FDA Database of glp1 Disproportionate Adverse EventsDocument20 pagesFEARS Database-FDA Database of glp1 Disproportionate Adverse EventsW Antonio Rivera MartínezNo ratings yet
- Cellular Aberration Lecture 1Document93 pagesCellular Aberration Lecture 1David Dwane Art SilorioNo ratings yet
- Maternal PhysiologyDocument73 pagesMaternal PhysiologyAbegail IbañezNo ratings yet
- Case Study Cerebrovascular AccidentDocument24 pagesCase Study Cerebrovascular Accidentdana86% (14)
- HSB Endocrine System and EyeDocument15 pagesHSB Endocrine System and EyeNisha JodhanNo ratings yet
- Metabolic ResponseDocument22 pagesMetabolic ResponsenelsonNo ratings yet
- Classification and Treatment of Bone TumorsDocument6 pagesClassification and Treatment of Bone TumorsRonald TejoprayitnoNo ratings yet
- Approach To Altered Mental StatusDocument19 pagesApproach To Altered Mental StatusMudassar SattarNo ratings yet
- Inflammation: Acute and ChronicDocument33 pagesInflammation: Acute and ChronicFort SalvadorNo ratings yet
- Abordaje AxilarDocument18 pagesAbordaje Axilarshoriwe68No ratings yet
- Radiology Semiotics of Diseases of Various SystemDocument7 pagesRadiology Semiotics of Diseases of Various Systemnikhil gendreNo ratings yet
- PSYC - 1001 - Chapter 4Document5 pagesPSYC - 1001 - Chapter 4Alaina PalmaNo ratings yet
- Physical Abuse in ChildrenDocument113 pagesPhysical Abuse in ChildrenDan Matthew BuriasNo ratings yet
- PCOME Ingram-Lopez Postmortem Examination ReportDocument8 pagesPCOME Ingram-Lopez Postmortem Examination ReportDylan SmithNo ratings yet
- Science Report - Stem CellsDocument7 pagesScience Report - Stem CellsKay NeethlingNo ratings yet
- HDFC Life Click 2 Protect Life - Product Brochure PDFDocument24 pagesHDFC Life Click 2 Protect Life - Product Brochure PDFvaibhav kumar KhokharNo ratings yet
- Medical Nursing Notes on Stress Response and Endocrine DisordersDocument37 pagesMedical Nursing Notes on Stress Response and Endocrine Disordersincognito100% (11)
- Biology June/July Paper: Application QuestionsDocument14 pagesBiology June/July Paper: Application QuestionsCHUA MING XIU HCINo ratings yet
- Week 8 Methods of Drug DeliveryDocument35 pagesWeek 8 Methods of Drug Deliveryejohannesen16No ratings yet
- Brain Edema and Disorders of CSF CirculationDocument68 pagesBrain Edema and Disorders of CSF CirculationDr. Yuvraj LahreNo ratings yet
- Constipation in Infants and Children - Evaluation - UpToDateDocument25 pagesConstipation in Infants and Children - Evaluation - UpToDatemonalisaNo ratings yet
- Thyroid SwellingDocument4 pagesThyroid SwellingShafiq ZahariNo ratings yet
- Liposuction Fat Embolism Syndrome DiagnosisDocument5 pagesLiposuction Fat Embolism Syndrome DiagnosisNgô Huỳnh ĐứcNo ratings yet
- CA Mammae Group Project on Breast CancerDocument16 pagesCA Mammae Group Project on Breast CancerNovita ningrumNo ratings yet
- Vertiginous EpilepsyDocument5 pagesVertiginous Epilepsyzudan2013No ratings yet
- Paediatrics & Biostatistics Notes - USMLE Step 2CKDocument102 pagesPaediatrics & Biostatistics Notes - USMLE Step 2CKDuncan89No ratings yet
- Cerebrospinal Fluid - AUBFDocument9 pagesCerebrospinal Fluid - AUBFMitch IbayNo ratings yet
- Week 4 A&P 2 Lab Exercise 27.1-27.14Document30 pagesWeek 4 A&P 2 Lab Exercise 27.1-27.14Rhaisa TsyboukovNo ratings yet
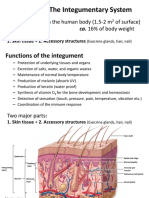
- BIOL 2210 Chapter 6, Integument System-1Document15 pagesBIOL 2210 Chapter 6, Integument System-1KellyPatrick SpencerNo ratings yet
- PATHFIT 1 Module 2 FinalDocument14 pagesPATHFIT 1 Module 2 FinalScarlet VillamorNo ratings yet