You might also like
- Basic Cell Theory - StructureDocument32 pagesBasic Cell Theory - StructureRuby SamarNo ratings yet
- Cell MembraneDocument29 pagesCell Membranesyraregencia15No ratings yet
- Cell MembraneDocument10 pagesCell Membraneharshika tembhurneNo ratings yet
- Chapter 4 Transport Across Cell MembranesDocument10 pagesChapter 4 Transport Across Cell Membranesnoreen dorani100% (1)
- Biology Notes: Topic-2ADocument9 pagesBiology Notes: Topic-2Alaiba amirNo ratings yet
- SF - MD.0145.T.22 Ally Omary MwambelaDocument10 pagesSF - MD.0145.T.22 Ally Omary MwambelaAlly OmaryNo ratings yet
- Cell MembraneDocument3 pagesCell MembraneQaran PrintingNo ratings yet
- 2structure and Functions of Cell MembraneDocument19 pages2structure and Functions of Cell MembraneedriansamaNo ratings yet
- Unit 3: How Are Eukaryotic Cells Organized Into Smaller Parts?Document6 pagesUnit 3: How Are Eukaryotic Cells Organized Into Smaller Parts?juan mondaNo ratings yet
- Question 1: Identify On A Cell Model or Diagram The Three Major Cell Regions (NucleusDocument10 pagesQuestion 1: Identify On A Cell Model or Diagram The Three Major Cell Regions (NucleusGracea Mae AranetaNo ratings yet
- Cell Membrane: Navigation SearchDocument10 pagesCell Membrane: Navigation SearchMuhassanah Abd AzizNo ratings yet
- 2a-Membranes and Transport 2Document67 pages2a-Membranes and Transport 2kfy8vr7wtwNo ratings yet
- Visual6 1 CellTransport-PassiveDocument21 pagesVisual6 1 CellTransport-PassiveLouise AndreiNo ratings yet
- Cell Structure and FucntionDocument14 pagesCell Structure and Fucntionashhkhan420No ratings yet
- Cell Membrane Transport ActivityDocument7 pagesCell Membrane Transport ActivityoctavianojanineNo ratings yet
- Cell MembraneDocument17 pagesCell MembraneEloisa MadrilenoNo ratings yet
- Explanation and Acceptable Reason YeahDocument3 pagesExplanation and Acceptable Reason YeahMj DeguzmanNo ratings yet
- The Cell MembraneDocument40 pagesThe Cell Membraneemmamarietaylor1597No ratings yet
- The Membranes of Different Types of Cells Are Involved in Many Different FunctionsDocument2 pagesThe Membranes of Different Types of Cells Are Involved in Many Different Functionssuperhigh06No ratings yet
- Cell Membrane: From Wikipedia, The Free EncyclopediaDocument9 pagesCell Membrane: From Wikipedia, The Free Encyclopediabanana_flavorNo ratings yet
- Cell MembraneDocument2 pagesCell MembraneZei-chen LocabenNo ratings yet
- Plasma Membrane: A Physical BarrierDocument3 pagesPlasma Membrane: A Physical BarrierDiane OrsuaNo ratings yet
- Cell Membrane SummaryDocument1 pageCell Membrane SummaryzafNo ratings yet
- General Functions and Characteristics: Cell MembraneDocument2 pagesGeneral Functions and Characteristics: Cell MembraneHannaNo ratings yet
- Full Download Human Anatomy 4th Edition Saladin Solutions ManualDocument35 pagesFull Download Human Anatomy 4th Edition Saladin Solutions Manualvocality.duettoqw603100% (36)
- Cell Membrane Function and StructureDocument6 pagesCell Membrane Function and Structurecris ibarraNo ratings yet
- Notes - Chapter 3 - Cellular Level of OrganizationDocument7 pagesNotes - Chapter 3 - Cellular Level of OrganizationRogie P. BacosaNo ratings yet
- Containing The Cell: Cell Membrane: Part I: Locating Physiology On The Web of KnowledgeDocument5 pagesContaining The Cell: Cell Membrane: Part I: Locating Physiology On The Web of KnowledgeEly TrajicoNo ratings yet
- Significance of Liquid in Biological Membranes and Transport MechanismDocument10 pagesSignificance of Liquid in Biological Membranes and Transport Mechanismroshan khanNo ratings yet
- This Learning Competency Is Extracted From The Deped Curriculum GuideDocument6 pagesThis Learning Competency Is Extracted From The Deped Curriculum GuideZuzuNo ratings yet
- Human Anatomy 4th Edition Saladin Solutions ManualDocument25 pagesHuman Anatomy 4th Edition Saladin Solutions ManualLeslieBuchanannjfy100% (52)
- Biology Module 2: Section 2.5Document20 pagesBiology Module 2: Section 2.5Nasteexo CaliNo ratings yet
- Cell Structure: Prepared by Dr. Sana AslamDocument41 pagesCell Structure: Prepared by Dr. Sana Aslamsajjad Ali balouch100% (1)
- Cell Structure and Function 1Document4 pagesCell Structure and Function 1Dianne LarozaNo ratings yet
- Cell Membrane: Dr. Samia ShahbazDocument26 pagesCell Membrane: Dr. Samia Shahbazreema aslamNo ratings yet
- Ch. 5 Pre-ReadingDocument7 pagesCh. 5 Pre-ReadingDJ ISAACS50% (2)
- 2.4 - Membranes: 2.4.1 - Draw and Label A Diagram To Show The Structure of MembranesDocument6 pages2.4 - Membranes: 2.4.1 - Draw and Label A Diagram To Show The Structure of MembranesDuong TongNo ratings yet
- Full Download Zoology 9th Edition Miller Solutions ManualDocument35 pagesFull Download Zoology 9th Edition Miller Solutions Manualcaveneywilliams100% (14)
- Laporan PhysioexDocument12 pagesLaporan Physioex12099964100% (1)
- Cell Structure - Cell Membrane and TransportDocument6 pagesCell Structure - Cell Membrane and TransportKulsoom UsmanNo ratings yet
- Cell Structures and FunctionsDocument5 pagesCell Structures and Functions9pjh6pzxmsNo ratings yet
- Cellular Transport Mechanism: General Biology 1Document12 pagesCellular Transport Mechanism: General Biology 1Kenn KennNo ratings yet
- Cell BiologyDocument3 pagesCell BiologyjamjamjamNo ratings yet
- WEEK5 ModuleLecDocument8 pagesWEEK5 ModuleLecMaricris GatdulaNo ratings yet
- (Notes) Cell MembraneDocument5 pages(Notes) Cell MembraneSiva Letchumi SelvanNo ratings yet
- Chapter 2 Student Copy ZoologyDocument5 pagesChapter 2 Student Copy Zoologyapi-530699294No ratings yet
- 3 - Plasma Membrane and Transport MechanismsDocument67 pages3 - Plasma Membrane and Transport MechanismsThom PaglinawanNo ratings yet
- Cell Membrane L3 CH3Document2 pagesCell Membrane L3 CH3Motie KassabNo ratings yet
- Chapter 2 Student Copy ZoologyDocument5 pagesChapter 2 Student Copy Zoologyapi-523871804No ratings yet
- Class NOTES - Cell Structures and Their Function.Document28 pagesClass NOTES - Cell Structures and Their Function.Cyril Josh MaisogNo ratings yet
- MODULE 12 - GB1 (Cell Transport)Document8 pagesMODULE 12 - GB1 (Cell Transport)NATHANIEL CORTEZNo ratings yet
- Dela Ysla Russell Iverson T. Biology1Document4 pagesDela Ysla Russell Iverson T. Biology1Elaiza Nicole AccatanNo ratings yet
- 1 2 3 4 5 6 MergedDocument90 pages1 2 3 4 5 6 MergedNAYEEMA JAMEEL ANUVANo ratings yet
- Membrane Structure and Function: Lecture OutlineDocument11 pagesMembrane Structure and Function: Lecture OutlineJohn Westly S. SabueroNo ratings yet
- 9gkneylgi4kueakdv8q9gkney50iscvv SolutionDocument2 pages9gkneylgi4kueakdv8q9gkney50iscvv SolutionArya MohajerNo ratings yet
- Animal Physiology Theory Lec1Document24 pagesAnimal Physiology Theory Lec1ao868598No ratings yet
- Components of Plasma Membranes: Learning ObjectivesDocument14 pagesComponents of Plasma Membranes: Learning ObjectivesAlthea Junsan BernabeNo ratings yet
- Chapter Overview: Subject Name The Cell Topic 1 Week 1Document9 pagesChapter Overview: Subject Name The Cell Topic 1 Week 1VeiliLookNo ratings yet
- Ecology - Lesson 1Document8 pagesEcology - Lesson 1LeighNo ratings yet
- Biophysics Also, It - Likewise, Biophysics Biophysics: TH THDocument4 pagesBiophysics Also, It - Likewise, Biophysics Biophysics: TH THLeighNo ratings yet
- GIBBS Free Energy, Module 5 Assignment QuizDocument1 pageGIBBS Free Energy, Module 5 Assignment QuizLeighNo ratings yet
- Module 7Document38 pagesModule 7LeighNo ratings yet
- Activity 1 Poster MakingDocument2 pagesActivity 1 Poster MakingLeighNo ratings yet
- Statistical Mechanics Gibbs Free EnergyDocument8 pagesStatistical Mechanics Gibbs Free EnergyLeighNo ratings yet
- Activity 1 FCDocument2 pagesActivity 1 FCLeighNo ratings yet
- Equilibrium KeyDocument4 pagesEquilibrium KeySunnyNo ratings yet
- Unit 3 (Module 6 - Biomolecules)Document8 pagesUnit 3 (Module 6 - Biomolecules)LeighNo ratings yet
- Module 8 ACTDocument6 pagesModule 8 ACTLeighNo ratings yet
- Module 9 ACTDocument3 pagesModule 9 ACTLeighNo ratings yet
- Equilibrium Practice Problems Answers 201314Document6 pagesEquilibrium Practice Problems Answers 201314Monceda Ramos MareinelleNo ratings yet
- Module 10 ACTDocument5 pagesModule 10 ACTLeighNo ratings yet
- Equilibrium Practice Problems Answers 201314Document6 pagesEquilibrium Practice Problems Answers 201314Monceda Ramos MareinelleNo ratings yet
- Equilibrium KeyDocument4 pagesEquilibrium KeySunnyNo ratings yet
- Fiction Books To Choose FromDocument15 pagesFiction Books To Choose FromLeighNo ratings yet
- Bio 3 BlabskeletonDocument12 pagesBio 3 BlabskeletonChucksters79No ratings yet
- Rizal's Life: Higher Education and Life AbroadDocument21 pagesRizal's Life: Higher Education and Life AbroadLeighNo ratings yet
- Evolution: Evidence and Mechanisms: Definitions: The Hypothesis ThatDocument28 pagesEvolution: Evidence and Mechanisms: Definitions: The Hypothesis ThatLeighNo ratings yet
- Rizal Was 16 Years Old When One Sunday in 1887 He Paid Visit To His Maternal Grandmother in TrozoDocument1 pageRizal Was 16 Years Old When One Sunday in 1887 He Paid Visit To His Maternal Grandmother in TrozoLeighNo ratings yet
- Common Name Scientific NameDocument2 pagesCommon Name Scientific NameLeighNo ratings yet
- Evolution: Evidence and Mechanisms: Definitions: The Hypothesis ThatDocument28 pagesEvolution: Evidence and Mechanisms: Definitions: The Hypothesis ThatLeighNo ratings yet
- Basketball Is A Team Sport Played by Two Teams of Five Players On A Rectangular CourtDocument13 pagesBasketball Is A Team Sport Played by Two Teams of Five Players On A Rectangular CourtLeighNo ratings yet
- Importance of Plant Anatomy in Agriculture1Document8 pagesImportance of Plant Anatomy in Agriculture1LeighNo ratings yet
- FriendshipDocument75 pagesFriendshipLeighNo ratings yet
- Psalms 23 The Lord Is My Shepherd Psalms 23 The Lord Is My ShepherdDocument5 pagesPsalms 23 The Lord Is My Shepherd Psalms 23 The Lord Is My ShepherdLeighNo ratings yet
- G G B M D: Enes and The Enetic Asis OF Etabolism AND EvelopmentDocument35 pagesG G B M D: Enes and The Enetic Asis OF Etabolism AND EvelopmentLeighNo ratings yet
- CH 12 Rate LawsDocument114 pagesCH 12 Rate LawsLeighNo ratings yet
- Leaves (Chapter 6)Document28 pagesLeaves (Chapter 6)LeighNo ratings yet
- Awwa C906-21Document14 pagesAwwa C906-21Juan PulgarNo ratings yet
- Opportunities and Barriers For Producing High Quality Fuels From The Pyrolysis of Scrap TiresDocument15 pagesOpportunities and Barriers For Producing High Quality Fuels From The Pyrolysis of Scrap TiresAbdisalamNo ratings yet
- Epocast 50a1-9816Document22 pagesEpocast 50a1-9816NICKYNo ratings yet
- Candle LabDocument5 pagesCandle LabHarrison Lee50% (2)
- Group CHM258 AssignmentDocument5 pagesGroup CHM258 Assignmentshark eyeNo ratings yet
- Topic 1 Manual 4th Edition Ver1 2023Document44 pagesTopic 1 Manual 4th Edition Ver1 2023Ha NaNo ratings yet
- 2C-03-01 Rev 0Document2 pages2C-03-01 Rev 0Christian Martínez G.No ratings yet
- Liquid Chromatography - Normal Phase: P Jandera, Univerzita Pardubice, Pardubice, Czech RepublicDocument12 pagesLiquid Chromatography - Normal Phase: P Jandera, Univerzita Pardubice, Pardubice, Czech RepublicEuderDiasNo ratings yet
- Ss 304 Microstructure ExplainedDocument5 pagesSs 304 Microstructure ExplainedNafillah AbdurrahmanNo ratings yet
- D and F BlockDocument20 pagesD and F BlockRuler HellNo ratings yet
- 2012MDEKEIJZERetal Indigocarmine Contributions-IICViennaDocument10 pages2012MDEKEIJZERetal Indigocarmine Contributions-IICViennaJhosua SimanullangNo ratings yet
- PND1000-3 Dryer ProductsDocument10 pagesPND1000-3 Dryer ProductsPartsGopher.comNo ratings yet
- Aniline PointDocument4 pagesAniline PointJalal OmarNo ratings yet
- BIO1 Lesson-12 PhotosynthesisDocument36 pagesBIO1 Lesson-12 PhotosynthesisCath Detoperez100% (1)
- Alcohol, Ether, Phenol WorksheetDocument22 pagesAlcohol, Ether, Phenol WorksheetSDMNo ratings yet
- 10 Wapda 2010Document140 pages10 Wapda 2010Sakhi RizviNo ratings yet
- Waterglass Vs AlkoxideDocument6 pagesWaterglass Vs AlkoxideShazni AhamedNo ratings yet
- Pharmacology For Dental and Allied Health SciencesDocument494 pagesPharmacology For Dental and Allied Health SciencesDevasmitha DevendraNo ratings yet
- NSF/ANSI/CAN 600 - 2021: Health Effects Evaluation and Criteria For Chemicals in Drinking WaterDocument201 pagesNSF/ANSI/CAN 600 - 2021: Health Effects Evaluation and Criteria For Chemicals in Drinking WaterFaiz SabianNo ratings yet
- Blow Mould DesignDocument78 pagesBlow Mould Designabhi858100% (13)
- PA SlideDocument36 pagesPA SlideRadhashyam GiriNo ratings yet
- Cv. NEW. FAJARDocument7 pagesCv. NEW. FAJARradenkobalarifinNo ratings yet
- Asphalt Safety and Best Practices: Table AC-1 of Cal/OSHA Code 5155Document7 pagesAsphalt Safety and Best Practices: Table AC-1 of Cal/OSHA Code 5155Olivia BugayNo ratings yet
- Piusi Air Operated P60 1 InstructionsDocument2 pagesPiusi Air Operated P60 1 Instructions673348300 Producto CartesNo ratings yet
- Health 9 Q2 FDocument42 pagesHealth 9 Q2 FVKNo ratings yet
- Worksheet 1Document16 pagesWorksheet 1All photos CloudNo ratings yet
- Standardisation of Herbal MedicinesDocument9 pagesStandardisation of Herbal MedicinesVinita GaonkarNo ratings yet
- Semi-Automatic Distillation Unit: Operating ManualDocument123 pagesSemi-Automatic Distillation Unit: Operating ManualNatalia EscobarNo ratings yet
- Potential Impact of Fall Armyworm On Wheat: Pest RiskDocument3 pagesPotential Impact of Fall Armyworm On Wheat: Pest RiskaiktiplarNo ratings yet
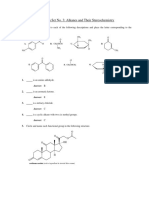
- Problem Set 3 - Alkanes and StereochemDocument6 pagesProblem Set 3 - Alkanes and StereochemKatrina Louise GonzalesNo ratings yet