You might also like
- Building Primitive Traps & SnaresDocument101 pagesBuilding Primitive Traps & SnaresJoseph Madr90% (10)
- Jialin Tian, Yabin Yu - Macaron Magic - 2011Document138 pagesJialin Tian, Yabin Yu - Macaron Magic - 2011Bogdan100% (1)
- Jialin Tian, Yabin Yu - Macaron Magic - 2011Document138 pagesJialin Tian, Yabin Yu - Macaron Magic - 2011Bogdan100% (1)
- Essene Moses Ten CommandmentsDocument4 pagesEssene Moses Ten Commandmentsjeryon91No ratings yet
- Request For Proposal Construction & Phase 1 OperationDocument116 pagesRequest For Proposal Construction & Phase 1 Operationsobhi100% (2)
- InteliLite AMF20-25Document2 pagesInteliLite AMF20-25albertooliveira100% (2)
- QP P1 APR 2023Document16 pagesQP P1 APR 2023Gil legaspiNo ratings yet
- Daily Assessment RecordDocument4 pagesDaily Assessment Recordapi-342236522100% (2)
- Nutraceuticals and Natural Product Derivatives: Disease Prevention & Drug DiscoveryFrom EverandNutraceuticals and Natural Product Derivatives: Disease Prevention & Drug DiscoveryNo ratings yet
- Compuestos Flavones FrutasDocument8 pagesCompuestos Flavones FrutasSuhey PérezNo ratings yet
- Anticancer Article 7Document14 pagesAnticancer Article 720-20311-014No ratings yet
- Analysis of Selected Phenolic Acids and Flavonoids in Amaranthus Cruentus and by HPLCDocument12 pagesAnalysis of Selected Phenolic Acids and Flavonoids in Amaranthus Cruentus and by HPLCWillam PariNo ratings yet
- Original Article Free Radical Scavenging Activities of Phytochemical Mixtures and Aqueous Methanolic Extracts Recovered From Processed Coffee LeavesDocument8 pagesOriginal Article Free Radical Scavenging Activities of Phytochemical Mixtures and Aqueous Methanolic Extracts Recovered From Processed Coffee LeavesHuy Trần Hoài NhậtNo ratings yet
- Total Phenolic and Flavonoid Contents and Antioxidant Activities of Two Raphanus Sativus L. Cultivars (Cherry Belle and Valentine)Document6 pagesTotal Phenolic and Flavonoid Contents and Antioxidant Activities of Two Raphanus Sativus L. Cultivars (Cherry Belle and Valentine)Almas TNo ratings yet
- Research Article: Correlations Between Antioxidant Activity and Alkaloids and Phenols of MacaDocument11 pagesResearch Article: Correlations Between Antioxidant Activity and Alkaloids and Phenols of MacaRebecca TiaraNo ratings yet
- In vitro antioxidant activity of Calamus rotang leaf and fruit extractsDocument5 pagesIn vitro antioxidant activity of Calamus rotang leaf and fruit extractsGanesh PrasaiNo ratings yet
- Cabbage Antioxidants Fight InflammationDocument6 pagesCabbage Antioxidants Fight InflammationReni WulansariNo ratings yet
- In Vitro Antioxidant Activity of Four Indian Medicinal PlantsDocument8 pagesIn Vitro Antioxidant Activity of Four Indian Medicinal Plantssaksae9747No ratings yet
- Antioksidan PhenolDocument5 pagesAntioksidan PhenoltridewantiwNo ratings yet
- Antioxidant Potency and Gcms Composition of Leaves of Artocarpus Altilis Park FosbDocument5 pagesAntioxidant Potency and Gcms Composition of Leaves of Artocarpus Altilis Park Fosbfitriani fajriNo ratings yet
- Tingkatan AntioksidanDocument5 pagesTingkatan AntioksidanMygameNo ratings yet
- Phenolic Composition and Antioxidant Properties of Some Traditionally Used Medicinal Plants Affected by The Extraction Time and HydrolysisDocument9 pagesPhenolic Composition and Antioxidant Properties of Some Traditionally Used Medicinal Plants Affected by The Extraction Time and HydrolysisBeatriz MelloNo ratings yet
- Season and Ecotype Effects On Soluble Phenolic Compounds Content and Antioxidant Potential of Tamarindus Indica and Mitragyna InermisDocument14 pagesSeason and Ecotype Effects On Soluble Phenolic Compounds Content and Antioxidant Potential of Tamarindus Indica and Mitragyna InermisSabrina JonesNo ratings yet
- Antioxidants and Chlorophyll in Cassava Leaves at Three Plant AgesDocument7 pagesAntioxidants and Chlorophyll in Cassava Leaves at Three Plant Agesdyan ayu puspariniNo ratings yet
- Wetland Plants High AntioxidantsDocument12 pagesWetland Plants High Antioxidantsvaishali shuklaNo ratings yet
- Investigation On The Antioxidant Activity of Leaves, Peels, Stems Bark, and Kernel of Mango (Mangifera Indica L.)Document4 pagesInvestigation On The Antioxidant Activity of Leaves, Peels, Stems Bark, and Kernel of Mango (Mangifera Indica L.)eliezer999No ratings yet
- Bioassay-Guided Isolation of Flavonoids From Caesalpinia Bonduc (L.) Roxb. and Evaluation of Their CytotoxicityDocument9 pagesBioassay-Guided Isolation of Flavonoids From Caesalpinia Bonduc (L.) Roxb. and Evaluation of Their Cytotoxicitynurul hidayatiNo ratings yet
- 2013 Oiirj CharyDocument8 pages2013 Oiirj CharyEstari MamidalaNo ratings yet
- (Acta Biologica Cracoviensia S. Botanica) DPPH Radical Scavenging Activity and Phenolic Compound Content in Different Leaf Extracts From Selected BLDocument7 pages(Acta Biologica Cracoviensia S. Botanica) DPPH Radical Scavenging Activity and Phenolic Compound Content in Different Leaf Extracts From Selected BLMonicaNo ratings yet
- Characterization of The Free Radical Scavenging Activity of Rose Hip Rosa Canina L ExtractDocument15 pagesCharacterization of The Free Radical Scavenging Activity of Rose Hip Rosa Canina L Extractriaulul IzmiNo ratings yet
- Improvement of Phenylpropanoid Production With Elicitor Treatments in Pimpinella Brachycarpa NakaiDocument16 pagesImprovement of Phenylpropanoid Production With Elicitor Treatments in Pimpinella Brachycarpa NakaiDinda Dwi SeptianiNo ratings yet
- Chemical Composition of Chrysanthemum Teas and Their Anti Inflammatory and Antioxidant PropertiesDocument9 pagesChemical Composition of Chrysanthemum Teas and Their Anti Inflammatory and Antioxidant PropertiesYou Jin LimNo ratings yet
- Anti-Cancer and Antioxidant Properties of Phenolics Isolated From Toona Sinensis A Juss Acetone Leaf ExtractDocument9 pagesAnti-Cancer and Antioxidant Properties of Phenolics Isolated From Toona Sinensis A Juss Acetone Leaf ExtractGrace Anastasia Ginting SinusingaNo ratings yet
- Antioxidant Activity of Mangrove Sonneratia Caseolaris L: Using The FRAP MethodDocument7 pagesAntioxidant Activity of Mangrove Sonneratia Caseolaris L: Using The FRAP MethodDita HarefaNo ratings yet
- Chen12-Flavonoid-Shift ReagentsDocument26 pagesChen12-Flavonoid-Shift ReagentsMarvinNo ratings yet
- Jung Et Al PDFDocument9 pagesJung Et Al PDFAri Syuhada PutraNo ratings yet
- Maity 2013Document8 pagesMaity 2013Sri LakshmiNo ratings yet
- PHCOG MAG.: Research Article in Vitro Antioxidant Activity of Asparagus Racemosus RootDocument8 pagesPHCOG MAG.: Research Article in Vitro Antioxidant Activity of Asparagus Racemosus RootmahemezNo ratings yet
- Antioxidative Activities of Water Extracts of Some Malaysian HerbsDocument8 pagesAntioxidative Activities of Water Extracts of Some Malaysian HerbsjivasumanaNo ratings yet
- ×piperita L. and Lavandula Angustifolia Mill. Extracts: Research Article MenthaDocument8 pages×piperita L. and Lavandula Angustifolia Mill. Extracts: Research Article MenthaWilayit Ali SaediNo ratings yet
- Lit CiteDocument8 pagesLit CitesimaNo ratings yet
- Total Antioxidant Power and Free RadicalDocument4 pagesTotal Antioxidant Power and Free RadicalDr. Varaprasad Bobbarala100% (1)
- Antioxidant and Antimicrobial Activity-M.maderaspatanaDocument9 pagesAntioxidant and Antimicrobial Activity-M.maderaspatanaAntoine PetrusNo ratings yet
- Accepted in Vitro Antioxidant Anti Inflammatory and Cytotoxicity Activities From Hexane Extract of Bryonopsis Laciniosa Fruits.Document15 pagesAccepted in Vitro Antioxidant Anti Inflammatory and Cytotoxicity Activities From Hexane Extract of Bryonopsis Laciniosa Fruits.umeshbt720No ratings yet
- Food and Chemical ToxicologyDocument7 pagesFood and Chemical ToxicologyEkajayanti KiningNo ratings yet
- Determination of Guava (Psidium Guajava L.) LeafDocument13 pagesDetermination of Guava (Psidium Guajava L.) LeafTiara Nugraeni EkaNo ratings yet
- Art2 V33No1p22-30Document9 pagesArt2 V33No1p22-30chanmiNo ratings yet
- Pol Is Aka RidDocument9 pagesPol Is Aka RidRiesa YositaNo ratings yet
- Research Journal of Pharmacy and Technology - PID - 2016-9-11-39 PDFDocument8 pagesResearch Journal of Pharmacy and Technology - PID - 2016-9-11-39 PDFAbd EL HakimNo ratings yet
- Flavonoids in Juglans Regia L. Leaves and Evaluation of in Vitro Antioxidant Activity Via Intracellular and Chemical MethodsDocument10 pagesFlavonoids in Juglans Regia L. Leaves and Evaluation of in Vitro Antioxidant Activity Via Intracellular and Chemical MethodsDyanne NegruNo ratings yet
- Article1429194127 - Zapata-Vahos Et AlDocument9 pagesArticle1429194127 - Zapata-Vahos Et AlVirnaIrdaPalupiNo ratings yet
- 05 Intan Soraya Che Sulaiman - Paling FunctionDocument14 pages05 Intan Soraya Che Sulaiman - Paling FunctionIdham ZaharudieNo ratings yet
- SampleDocument6 pagesSamplejayven minguillanNo ratings yet
- Antioxidant Activities of Methanolic and Aqueous Extracts From Leaves of Martynia Annua Linn.Document10 pagesAntioxidant Activities of Methanolic and Aqueous Extracts From Leaves of Martynia Annua Linn.Kseniia RatushnaNo ratings yet
- Phytochemical and Antioxidant Studies On Leaf Extracts of Muntingia Calabura LDocument6 pagesPhytochemical and Antioxidant Studies On Leaf Extracts of Muntingia Calabura LAlwyn James AgcongayNo ratings yet
- Antibacterial and Antioxidant Activity of Parmotrema Reticulatum Obtained F PDFDocument5 pagesAntibacterial and Antioxidant Activity of Parmotrema Reticulatum Obtained F PDFIrin TandelNo ratings yet
- Phytochemicals, Antioxidant Properties and Anticancer Investigations of The Different Parts of Several Gingers Species (Document6 pagesPhytochemicals, Antioxidant Properties and Anticancer Investigations of The Different Parts of Several Gingers Species (vanessaNo ratings yet
- Some Contained-Allantoin Plants: In-Vitro Antioxidant Activities of The Ethanolic Extracts ofDocument14 pagesSome Contained-Allantoin Plants: In-Vitro Antioxidant Activities of The Ethanolic Extracts ofmy nameNo ratings yet
- Evaluation of Antioxidant Activity of Clove Syzygium AromaticumDocument8 pagesEvaluation of Antioxidant Activity of Clove Syzygium AromaticumTrina ViskhawatNo ratings yet
- Aloe VeraDocument0 pagesAloe VeraSyahrul FarhanahNo ratings yet
- Antioxidant Activities of Methanol and Dichloromethane Blend Extracts of Caesalpinia Volkensii Harms. and Carissa Edulis (Forssk.) in VitroDocument8 pagesAntioxidant Activities of Methanol and Dichloromethane Blend Extracts of Caesalpinia Volkensii Harms. and Carissa Edulis (Forssk.) in VitroInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- 5c0a PDFDocument9 pages5c0a PDFnelisaNo ratings yet
- ORAC and CancerDocument8 pagesORAC and CancerPaolo Robert BuenoNo ratings yet
- 2 IFRJ 20 (03) 2013 Asmah PDFDocument6 pages2 IFRJ 20 (03) 2013 Asmah PDFKarina Winda BessufiNo ratings yet
- 1 s2.0 S2307410823000524 MainDocument4 pages1 s2.0 S2307410823000524 MainTanti 01No ratings yet
- Hasmila 2019 J. Phys. Conf. Ser. 1341 032027Document7 pagesHasmila 2019 J. Phys. Conf. Ser. 1341 032027Rebeca MaciasNo ratings yet
- Antioxidant Phenolic Constituents in Roots of Rheum OfficinaDocument7 pagesAntioxidant Phenolic Constituents in Roots of Rheum OfficinaIman IltihamNo ratings yet
- Essential Oils From Eucalyptus StaigerianaDocument10 pagesEssential Oils From Eucalyptus Staigerianaasa chiNo ratings yet
- Αntioxidant Activity of Cynara Scolymus L. AndDocument6 pagesΑntioxidant Activity of Cynara Scolymus L. Andasa chiNo ratings yet
- Metabolomics Analysis To Evaluate The AntibacterialDocument10 pagesMetabolomics Analysis To Evaluate The Antibacterialasa chiNo ratings yet
- Αntioxidant Activity of Cynara Scolymus L. AndDocument6 pagesΑntioxidant Activity of Cynara Scolymus L. Andasa chiNo ratings yet
- Metabolomics Analysis To Evaluate The AntibacterialDocument10 pagesMetabolomics Analysis To Evaluate The Antibacterialasa chiNo ratings yet
- Anti-Tyrosinase and Antioxidant ActivityDocument18 pagesAnti-Tyrosinase and Antioxidant Activityasa chiNo ratings yet
- Nanotechnology For Plant Disease Management 2Document24 pagesNanotechnology For Plant Disease Management 2asa chiNo ratings yet
- Nanotechnology For Plant Disease Management 2Document24 pagesNanotechnology For Plant Disease Management 2asa chiNo ratings yet
- Essential Oils From Eucalyptus StaigerianaDocument10 pagesEssential Oils From Eucalyptus Staigerianaasa chiNo ratings yet
- Antifungal Activity of Michelia AlbaDocument25 pagesAntifungal Activity of Michelia Albaasa chiNo ratings yet
- Antifungal and AntiaflatoxigenicDocument10 pagesAntifungal and Antiaflatoxigenicasa chiNo ratings yet
- Antifungal and AntiaflatoxigenicDocument10 pagesAntifungal and Antiaflatoxigenicasa chiNo ratings yet
- Anti-Diabetic Activity of Crude Pistacia LentiscusDocument6 pagesAnti-Diabetic Activity of Crude Pistacia Lentiscusasa chiNo ratings yet
- Anti-Diabetic Activity of Crude Pistacia LentiscusDocument6 pagesAnti-Diabetic Activity of Crude Pistacia Lentiscusasa chiNo ratings yet
- 18.food and Medicinal Values of Roselle Hibiscus Sabdariffa L Linne Malvaceae Plant Parts A Review 121Document7 pages18.food and Medicinal Values of Roselle Hibiscus Sabdariffa L Linne Malvaceae Plant Parts A Review 121asa chiNo ratings yet
- 18.food and Medicinal Values of Roselle Hibiscus Sabdariffa L Linne Malvaceae Plant Parts A Review 121Document7 pages18.food and Medicinal Values of Roselle Hibiscus Sabdariffa L Linne Malvaceae Plant Parts A Review 121asa chiNo ratings yet
- Antifungal Activity of Michelia AlbaDocument25 pagesAntifungal Activity of Michelia Albaasa chiNo ratings yet
- An Ethnomedicinal SurveyDocument10 pagesAn Ethnomedicinal Surveyasa chiNo ratings yet
- EUCAST definitions clinical breakpoints epidemiological cut-off valuesDocument1 pageEUCAST definitions clinical breakpoints epidemiological cut-off valuesMarcelo UGNo ratings yet
- Ait Ouazzou2011Document9 pagesAit Ouazzou2011asa chiNo ratings yet
- Ait Ouazzou2011Document9 pagesAit Ouazzou2011asa chiNo ratings yet
- Chemical Composition Antimicrobial AntioDocument8 pagesChemical Composition Antimicrobial AntioDanaNo ratings yet
- Toxicity of Mentha piperita oil against pestsDocument8 pagesToxicity of Mentha piperita oil against pestsasa chiNo ratings yet
- Antifungal Activity of ChitosanDocument13 pagesAntifungal Activity of Chitosanasa chiNo ratings yet
- Algicidal Properties of Extracts From CinnamomumDocument10 pagesAlgicidal Properties of Extracts From Cinnamomumasa chiNo ratings yet
- Acaricidal Activity of CompoundsDocument11 pagesAcaricidal Activity of Compoundsasa chiNo ratings yet
- Alkaloids of The Genus Cinnamomum Isolation Structure and Synthesis of Cinnamolaurine 1969 Tetrahedron LettersDocument4 pagesAlkaloids of The Genus Cinnamomum Isolation Structure and Synthesis of Cinnamolaurine 1969 Tetrahedron Lettersasa chiNo ratings yet
- IPO Fact Sheet - Accordia Golf Trust 140723Document4 pagesIPO Fact Sheet - Accordia Golf Trust 140723Invest StockNo ratings yet
- 99th Indian Science Congress (Bhubaneshwar)Document95 pages99th Indian Science Congress (Bhubaneshwar)Aadarsh DasNo ratings yet
- I Button Proper TDocument4 pagesI Button Proper TmariammariaNo ratings yet
- Managing Post COVID SyndromeDocument4 pagesManaging Post COVID SyndromeReddy VaniNo ratings yet
- 41 PDFsam Redis CookbookDocument5 pages41 PDFsam Redis CookbookHữu Hưởng NguyễnNo ratings yet
- Structural IfpDocument4 pagesStructural IfpDanny NguyenNo ratings yet
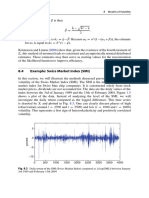
- 8.4 Example: Swiss Market Index (SMI) : 188 8 Models of VolatilityDocument3 pages8.4 Example: Swiss Market Index (SMI) : 188 8 Models of VolatilityNickesh ShahNo ratings yet
- Internal Resistance and Matching in Voltage SourceDocument8 pagesInternal Resistance and Matching in Voltage SourceAsif Rasheed Rajput100% (1)
- How Do I Prepare For Public Administration For IAS by Myself Without Any Coaching? Which Books Should I Follow?Document3 pagesHow Do I Prepare For Public Administration For IAS by Myself Without Any Coaching? Which Books Should I Follow?saiviswanath0990100% (1)
- EST I - Literacy Test I (Language)Document20 pagesEST I - Literacy Test I (Language)Mohammed Abdallah100% (1)
- A61C00100 Communication and Employee Engagement by Mary Welsh 2Document19 pagesA61C00100 Communication and Employee Engagement by Mary Welsh 2Moeshfieq WilliamsNo ratings yet
- Time Table Semester II 2019-2020Document18 pagesTime Table Semester II 2019-2020Shiv Kumar MeenaNo ratings yet
- Virtio-Fs - A Shared File System For Virtual MachinesDocument21 pagesVirtio-Fs - A Shared File System For Virtual MachinesLeseldelaterreNo ratings yet
- College of Physical Therapy Produces Skilled ProfessionalsDocument6 pagesCollege of Physical Therapy Produces Skilled ProfessionalsRia Mae Abellar SalvadorNo ratings yet
- Abbreviation Meaning Notes: Cibo ("With Food")Document4 pagesAbbreviation Meaning Notes: Cibo ("With Food")TantriNo ratings yet
- Lesson Plan 2 Revised - Morgan LegrandDocument19 pagesLesson Plan 2 Revised - Morgan Legrandapi-540805523No ratings yet
- Project Wise CSR Expenditure FY202223Document15 pagesProject Wise CSR Expenditure FY202223gowowor677No ratings yet
- Visualization BenchmarkingDocument15 pagesVisualization BenchmarkingRanjith S100% (1)
- Presentation of Urban RegenerationsDocument23 pagesPresentation of Urban RegenerationsRafiuddin RoslanNo ratings yet
- Grace Lipsini1 2 3Document4 pagesGrace Lipsini1 2 3api-548923370No ratings yet
- 1491559890CL 0417 PDFDocument48 pages1491559890CL 0417 PDFCoolerAdsNo ratings yet
- GRP Product CatalogueDocument57 pagesGRP Product CatalogueMulyana alcNo ratings yet
- Maths SolutionDocument10 pagesMaths SolutionAbhay KumarNo ratings yet
- Staining TechniquesDocument31 pagesStaining TechniquesKhadija JaraNo ratings yet