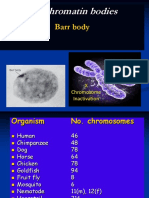
You might also like
- Mendelian Inheritance in Man: Catalogs of Autosomal Dominant, Autosomal Recessive, and X-Linked PhenotypesFrom EverandMendelian Inheritance in Man: Catalogs of Autosomal Dominant, Autosomal Recessive, and X-Linked PhenotypesNo ratings yet
- FullDocument5 pagesFull20100029No ratings yet
- 2011 Koumbaris HumMolGenetDocument12 pages2011 Koumbaris HumMolGenetEduardo PasteneNo ratings yet
- Detection of Pericentric Inversion of X Chromosome in A Male Fetus (American Journal of Medical Genetics, Vol. 87, Issue 4) (1999)Document3 pagesDetection of Pericentric Inversion of X Chromosome in A Male Fetus (American Journal of Medical Genetics, Vol. 87, Issue 4) (1999)Araceli Enríquez OvandoNo ratings yet
- Nama: Dewi Fatmawati NPM: 1814141016 Produksi Ternak: Genes Are Located On ChromosomesDocument8 pagesNama: Dewi Fatmawati NPM: 1814141016 Produksi Ternak: Genes Are Located On ChromosomesDewi FatmawatiNo ratings yet
- Sex Chromosome Aberrations and Transsexualism: VOL. 79, NO. 3, MARCH 2003Document2 pagesSex Chromosome Aberrations and Transsexualism: VOL. 79, NO. 3, MARCH 2003Fvg Fvg FvgNo ratings yet
- Genetic LinkageDocument82 pagesGenetic LinkagesahiddinpvictoryNo ratings yet
- Sect 15 Linkage&RecombinationDocument37 pagesSect 15 Linkage&RecombinationUsnia BaiqNo ratings yet
- 1 s2.0 S2666379124000636 MainDocument8 pages1 s2.0 S2666379124000636 MainXusetaSanzNo ratings yet
- Sect15Linkage&Recombination PDFDocument38 pagesSect15Linkage&Recombination PDFRauven Jean Erodias FadriquelaNo ratings yet
- Chromosome Imbalance As A Driver of Sex Disparity in DiseaseDocument12 pagesChromosome Imbalance As A Driver of Sex Disparity in DiseaseFirmanNo ratings yet
- 143 FullDocument8 pages143 FullDokdem AjaNo ratings yet
- Crossover Interference Underlies Sex Differences in Recombination RatesDocument4 pagesCrossover Interference Underlies Sex Differences in Recombination RatesAlfiaNo ratings yet
- 4.1. Chromosomes, Genes, Alleles and Mutations: Topic 10 & Topic 4: GeneticsDocument14 pages4.1. Chromosomes, Genes, Alleles and Mutations: Topic 10 & Topic 4: GeneticsMorgan Locke50% (2)
- Origin and Evolution of X Chromosome Inactivation: Joost Gribnau and J Anton GrootegoedDocument8 pagesOrigin and Evolution of X Chromosome Inactivation: Joost Gribnau and J Anton GrootegoedLuz MichueNo ratings yet
- Drosophzla Melanogasterlj: David T. Suzuki, David Baillie Dilys Parry3Document12 pagesDrosophzla Melanogasterlj: David T. Suzuki, David Baillie Dilys Parry3vimalNo ratings yet
- Fate of SRY, PABY, DYS1, DYZ3 and DYZ1 Loci in Indian Patients Harbouring Sex Chromosomal AnomaliesDocument11 pagesFate of SRY, PABY, DYS1, DYZ3 and DYZ1 Loci in Indian Patients Harbouring Sex Chromosomal Anomaliesrenny100% (1)
- 1990 BreeuwerWerr MicroorgChromDestructionDocument3 pages1990 BreeuwerWerr MicroorgChromDestructionIsworo RukmiNo ratings yet
- Forensic Science International: Genetics Supplement SeriesDocument3 pagesForensic Science International: Genetics Supplement SeriesAdrienn MatheNo ratings yet
- Ovarian Fibromas and Cystadenofibromas: Features of The Fibrous ComponentDocument7 pagesOvarian Fibromas and Cystadenofibromas: Features of The Fibrous ComponentSandu AlexandraNo ratings yet
- Ionasescu 1996Document6 pagesIonasescu 1996Ali AmokraneNo ratings yet
- Margarit 2000Document4 pagesMargarit 2000dad dzd adaNo ratings yet
- Molecular Cytogenetic Analysis of A DuplDocument7 pagesMolecular Cytogenetic Analysis of A DuplgeorgianaNo ratings yet
- Pnas00101 0183 PDFDocument4 pagesPnas00101 0183 PDFGAMEPLAY & FUNNY MOMENTSNo ratings yet
- Sex Origin VertebratesDocument9 pagesSex Origin VertebratesBiologia MencionNo ratings yet
- Chromosomal Basis of Inheritance - Final PDF-1Document3 pagesChromosomal Basis of Inheritance - Final PDF-1Aishwarya ShuklaNo ratings yet
- Chapter 3Document7 pagesChapter 3candra mahardikaNo ratings yet
- XP Deletions Associated With Autism in Three FemalesDocument6 pagesXP Deletions Associated With Autism in Three FemalesRami NaNo ratings yet
- Robertsonian TranslocationDocument10 pagesRobertsonian TranslocationJan ChenNo ratings yet
- The Human Y Chromosome Comes of AgeDocument15 pagesThe Human Y Chromosome Comes of Agegoldspotter9841No ratings yet
- 5 Pages Evidence of Fetal Microchimerism in Hashimotos ThyroiditisDocument5 pages5 Pages Evidence of Fetal Microchimerism in Hashimotos ThyroiditisKristinDaigleNo ratings yet
- Early X Chromosome Inactivation During Human Preimplantation Development Revealed by Single-Cell RNA-sequencingDocument12 pagesEarly X Chromosome Inactivation During Human Preimplantation Development Revealed by Single-Cell RNA-sequencinglizaNo ratings yet
- GenesDocument15 pagesGenes6 ANo ratings yet
- Chatterjee2017 Article DosageCompensationAndItsRolesIDocument19 pagesChatterjee2017 Article DosageCompensationAndItsRolesIAngelicaNo ratings yet
- Sex Chromatin Bodies: Barr BodyDocument45 pagesSex Chromatin Bodies: Barr BodyKhaleelMetlakNo ratings yet
- A Career in Chromosomes: Matters! Matters!Document1 pageA Career in Chromosomes: Matters! Matters!Hartford CourantNo ratings yet
- Chapter 12Document40 pagesChapter 12Sathish KumarNo ratings yet
- Clock Mutants of Drosophila: MelanogasterDocument5 pagesClock Mutants of Drosophila: MelanogasterfranciscoNo ratings yet
- Handout - Sex DeterminationDocument33 pagesHandout - Sex DeterminationAyah AbusafiaNo ratings yet
- 5 & 6 Genetics 2021Document116 pages5 & 6 Genetics 2021Greeshma ReddyNo ratings yet
- HeredityDocument39 pagesHereditySamantha CunananNo ratings yet
- Molecular Support For Heterogonesis Resulting in Sesquizygotic TwinningDocument8 pagesMolecular Support For Heterogonesis Resulting in Sesquizygotic TwinningartNo ratings yet
- Papers: Fetal Nuchal Translucency: Ultrasound Screening For Chromosomal Defects in First Trimester ofDocument3 pagesPapers: Fetal Nuchal Translucency: Ultrasound Screening For Chromosomal Defects in First Trimester ofHao Keat HoeNo ratings yet
- Amstutz Ajnr 2006 27:794Document5 pagesAmstutz Ajnr 2006 27:794jeheiserman1No ratings yet
- 1 s2.0 S0960982213010270 MainDocument3 pages1 s2.0 S0960982213010270 MainPiyaaNo ratings yet
- Twins With Schizophrenia: Genes or Germs?: by James O. Davis and Jeanne A. PhelpsDocument6 pagesTwins With Schizophrenia: Genes or Germs?: by James O. Davis and Jeanne A. PhelpsAdrienn MatheNo ratings yet
- Meta Gene: Mohit Kumar, Vandana Lal, Shilpa Chapadgaonkar, Saurabh Kumar BhattacharyaDocument6 pagesMeta Gene: Mohit Kumar, Vandana Lal, Shilpa Chapadgaonkar, Saurabh Kumar BhattacharyafaeNo ratings yet
- Development and Targeted Application of A Rapid QF-PCR Test For Sex Chromosome ImbalanceDocument10 pagesDevelopment and Targeted Application of A Rapid QF-PCR Test For Sex Chromosome ImbalancerennyNo ratings yet
- 트트트Document7 pages트트트100 568 최재훈No ratings yet
- Molecular Biology of Turner's Syndrome-Archive of Disease in ChildhoodDocument2 pagesMolecular Biology of Turner's Syndrome-Archive of Disease in ChildhoodNinaNo ratings yet
- Allison Et Al. - 1952 - Blood Groups in Some East African TribesDocument8 pagesAllison Et Al. - 1952 - Blood Groups in Some East African TribesSalam SawadogoNo ratings yet
- ChiasmataDocument4 pagesChiasmataDibyakNo ratings yet
- Mucoceles of The Paranasal Sinuses:: MR Imaging With CT CorrelationDocument6 pagesMucoceles of The Paranasal Sinuses:: MR Imaging With CT CorrelationArifah GulamanisNo ratings yet
- Babcock Effects of Differential Selection in The Sexes On Cytonuclear DynamicsDocument16 pagesBabcock Effects of Differential Selection in The Sexes On Cytonuclear Dynamicsmilorad miliNo ratings yet
- NOTES - CH 15 - Sex Determination - Linkage - SlideshowDocument59 pagesNOTES - CH 15 - Sex Determination - Linkage - Slideshowkaren milloNo ratings yet
- Characterization by Fluorescence and Electron Microscopy in Situ Hybridization of A Double Y IsochromosomeDocument4 pagesCharacterization by Fluorescence and Electron Microscopy in Situ Hybridization of A Double Y IsochromosomeEloy Guillermo Mijares EchezuríaNo ratings yet
- Genetically Homozygous Choriocarcinoma Following Hydatidiform MoleDocument5 pagesGenetically Homozygous Choriocarcinoma Following Hydatidiform MoleFeby HidasariNo ratings yet
- Oral Session I Which Theory Do The Structural Brain Changes Support?Document2 pagesOral Session I Which Theory Do The Structural Brain Changes Support?antonioopNo ratings yet
- Battaglia 9 25 14Document81 pagesBattaglia 9 25 14Bravo BNo ratings yet
- Ancient Sumer Flip BookDocument9 pagesAncient Sumer Flip Bookapi-198624210No ratings yet
- Cipet Bhubaneswar Skill Development CoursesDocument1 pageCipet Bhubaneswar Skill Development CoursesDivakar PanigrahiNo ratings yet
- Microfinance Ass 1Document15 pagesMicrofinance Ass 1Willard MusengeyiNo ratings yet
- 21 Tara Mantra-Wps OfficeDocument25 pages21 Tara Mantra-Wps OfficeAlteo FallaNo ratings yet
- Natural Cataclysms and Global ProblemsDocument622 pagesNatural Cataclysms and Global ProblemsphphdNo ratings yet
- Week - 2 Lab - 1 - Part I Lab Aim: Basic Programming Concepts, Python InstallationDocument13 pagesWeek - 2 Lab - 1 - Part I Lab Aim: Basic Programming Concepts, Python InstallationSahil Shah100% (1)
- Dynalift Sed0804679lDocument1 pageDynalift Sed0804679lzaryab khanNo ratings yet
- Fire Prevention Plan Template - FINAL 5-30-08Document5 pagesFire Prevention Plan Template - FINAL 5-30-08Peter GeorgeNo ratings yet
- EP001 LifeCoachSchoolTranscriptDocument13 pagesEP001 LifeCoachSchoolTranscriptVan GuedesNo ratings yet
- Strategic Marketing: The University of Lahore (Islamabad Campus)Document3 pagesStrategic Marketing: The University of Lahore (Islamabad Campus)Sumaira IrshadNo ratings yet
- Angel C. Delos Santos: Personal DataDocument8 pagesAngel C. Delos Santos: Personal DataAngel Cascayan Delos SantosNo ratings yet
- Data Network Unit 6 - UCDocument15 pagesData Network Unit 6 - UCANISHA DONDENo ratings yet
- Bossypants Autobiography and Womens SelvesDocument26 pagesBossypants Autobiography and Womens SelvesCamila Paz GutiérrezNo ratings yet
- EZ Water Calculator 3.0.2Document4 pagesEZ Water Calculator 3.0.2adriano70No ratings yet
- MolnarDocument8 pagesMolnarMaDzik MaDzikowskaNo ratings yet
- Marion Nicoll: Life & Work by Catharine MastinDocument147 pagesMarion Nicoll: Life & Work by Catharine MastinArt Canada InstituteNo ratings yet
- Unit 2 - Industrial Engineering & Ergonomics - WWW - Rgpvnotes.inDocument15 pagesUnit 2 - Industrial Engineering & Ergonomics - WWW - Rgpvnotes.inSACHIN HANAGALNo ratings yet
- Albert Roussel, Paul LandormyDocument18 pagesAlbert Roussel, Paul Landormymmarriuss7No ratings yet
- Inventions Over The Last 100 YearsDocument3 pagesInventions Over The Last 100 YearsHombreMorado GamerYTNo ratings yet
- CCDocument5 pagesCCnazmulNo ratings yet
- Guideline - Research ProposalDocument38 pagesGuideline - Research ProposalRASNo ratings yet
- Omnitron CatalogDocument180 pagesOmnitron Catalogjamal AlawsuNo ratings yet
- Lesson 1: Composition: Parts of An EggDocument22 pagesLesson 1: Composition: Parts of An Eggjohn michael pagalaNo ratings yet
- 4B - Urp - Shavya's FarmDocument22 pages4B - Urp - Shavya's FarmSnehansh KishoreNo ratings yet
- Liquitex Soft Body BookletDocument12 pagesLiquitex Soft Body Booklethello belloNo ratings yet
- Cpar Characteristics and Functions Week 3Document128 pagesCpar Characteristics and Functions Week 3christianwood0117No ratings yet
- Internship Format HRMI620Document4 pagesInternship Format HRMI620nimra tariqNo ratings yet
- Chief Complaint: History TakingDocument9 pagesChief Complaint: History TakingMohamad ZulfikarNo ratings yet
- Reflection On The PoorDocument5 pagesReflection On The Poorapi-347831792No ratings yet
- Contemporary Philippine Arts From The Regions: Quarter 1Document11 pagesContemporary Philippine Arts From The Regions: Quarter 1JUN GERONANo ratings yet
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeFrom EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeRating: 2 out of 5 stars2/5 (1)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsFrom EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNo ratings yet
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisFrom EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisRating: 4.5 out of 5 stars4.5/5 (42)
- The Age of Magical Overthinking: Notes on Modern IrrationalityFrom EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityRating: 4 out of 5 stars4/5 (24)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedRating: 5 out of 5 stars5/5 (80)
- The Obesity Code: Unlocking the Secrets of Weight LossFrom EverandThe Obesity Code: Unlocking the Secrets of Weight LossRating: 4 out of 5 stars4/5 (5)
- Summary: The Myth of Normal: Trauma, Illness, and Healing in a Toxic Culture By Gabor Maté MD & Daniel Maté: Key Takeaways, Summary & AnalysisFrom EverandSummary: The Myth of Normal: Trauma, Illness, and Healing in a Toxic Culture By Gabor Maté MD & Daniel Maté: Key Takeaways, Summary & AnalysisRating: 4 out of 5 stars4/5 (9)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsFrom EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsRating: 5 out of 5 stars5/5 (1)
- Sleep Stories for Adults: Overcome Insomnia and Find a Peaceful AwakeningFrom EverandSleep Stories for Adults: Overcome Insomnia and Find a Peaceful AwakeningRating: 4 out of 5 stars4/5 (3)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaFrom EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaRating: 4.5 out of 5 stars4.5/5 (266)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDFrom EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDRating: 5 out of 5 stars5/5 (1)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsFrom EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsRating: 3.5 out of 5 stars3.5/5 (3)
- Outlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisFrom EverandOutlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisRating: 4 out of 5 stars4/5 (1)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessFrom EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessRating: 4.5 out of 5 stars4.5/5 (328)
- The Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeFrom EverandThe Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeRating: 4.5 out of 5 stars4.5/5 (253)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsFrom EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsRating: 4.5 out of 5 stars4.5/5 (169)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.From EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Rating: 4.5 out of 5 stars4.5/5 (110)
- To Explain the World: The Discovery of Modern ScienceFrom EverandTo Explain the World: The Discovery of Modern ScienceRating: 3.5 out of 5 stars3.5/5 (51)
- An Autobiography of Trauma: A Healing JourneyFrom EverandAn Autobiography of Trauma: A Healing JourneyRating: 5 out of 5 stars5/5 (2)
- 12 Rules for Life by Jordan B. Peterson - Book Summary: An Antidote to ChaosFrom Everand12 Rules for Life by Jordan B. Peterson - Book Summary: An Antidote to ChaosRating: 4.5 out of 5 stars4.5/5 (207)