You might also like
- Sketchy Micro Table of Contents - MicroDocument5 pagesSketchy Micro Table of Contents - MicromarandrNo ratings yet
- Rodaks 6th Ed. Hema Lab Midterm ReviewerDocument30 pagesRodaks 6th Ed. Hema Lab Midterm ReviewerGerald SorianoNo ratings yet
- Central DogmaDocument4 pagesCentral DogmaRodriguez MiaNo ratings yet
- 2012-Ch 26 H. 627-Integrated Pest Management and Pest Control Current and Future TreatmentDocument682 pages2012-Ch 26 H. 627-Integrated Pest Management and Pest Control Current and Future TreatmentDwiNo ratings yet
- Do GargaloDocument9 pagesDo GargaloGustavo FelpeNo ratings yet
- The Infuence of Culture-Dependent NativeDocument14 pagesThe Infuence of Culture-Dependent NativeAfraNo ratings yet
- LT I> Anopheles Aquasalis&lDocument15 pagesLT I> Anopheles Aquasalis&lDwiNo ratings yet
- Malaria Transmission After Five Years of Vector Control On Bioko Island, Equatorial GuineaDocument14 pagesMalaria Transmission After Five Years of Vector Control On Bioko Island, Equatorial GuineaLSEGURA1212No ratings yet
- Molecular Research On Plasmodium Infection and ImmunityDocument3 pagesMolecular Research On Plasmodium Infection and ImmunityJean Bosco MBONIMPANo ratings yet
- Anopheles Gambiae S.S. Plasmodium FalciparumDocument9 pagesAnopheles Gambiae S.S. Plasmodium Falciparumibrahima1968No ratings yet
- vms3 434Document9 pagesvms3 434ali sherNo ratings yet
- Artigo Cientifico EpidemiologiaDocument10 pagesArtigo Cientifico Epidemiologiaroooberta3No ratings yet
- Research Interests For WebDocument2 pagesResearch Interests For WebMBAOLDNo ratings yet
- Streptococcus PyogenesDocument11 pagesStreptococcus PyogenesverajovvanaNo ratings yet
- Infectious Diseases 2Document17 pagesInfectious Diseases 2JohnchubyNo ratings yet
- s12916 022 02597 6Document12 pagess12916 022 02597 6Darshan GandhiNo ratings yet
- Larval Predation in Malaria Vectors and Its Potential Implication in Malaria Transmission: An Overlooked Ecosystem Service?Document11 pagesLarval Predation in Malaria Vectors and Its Potential Implication in Malaria Transmission: An Overlooked Ecosystem Service?umar yusufNo ratings yet
- Parasita No Intestino Do MosquitoDocument8 pagesParasita No Intestino Do MosquitoGustavo FelpeNo ratings yet
- 02 Outdoor Biting by Anopheles MosquitoesDocument8 pages02 Outdoor Biting by Anopheles MosquitoesDaniel VargasNo ratings yet
- Pathogens 11 00811Document5 pagesPathogens 11 00811octav drutaNo ratings yet
- Genetic Approaches For Malaria Control: Marcelo Jacobs-LorenaDocument14 pagesGenetic Approaches For Malaria Control: Marcelo Jacobs-Lorenayenniffer VicenteNo ratings yet
- Plasmodium and Other Malaria-Like Parasites. A Neglected CharacteristicDocument3 pagesPlasmodium and Other Malaria-Like Parasites. A Neglected CharacteristicMohammed KouidriNo ratings yet
- Evaluation of The Evolutionary Genetics and Population Structure of Culex Pipiens On Knockdown Resistance (KDR) Mutations and The Mtdna Coi GeneDocument14 pagesEvaluation of The Evolutionary Genetics and Population Structure of Culex Pipiens On Knockdown Resistance (KDR) Mutations and The Mtdna Coi GeneLuize Hoffmann Dall'AgneseNo ratings yet
- Ni Hms 747150Document26 pagesNi Hms 747150nydiacastillom2268No ratings yet
- Giardia Duodenalis in Patients With Diarrhea and Various Animals in Northeastern ChinaDocument11 pagesGiardia Duodenalis in Patients With Diarrhea and Various Animals in Northeastern ChinaFlávia UchôaNo ratings yet
- Fimmu 13 848267Document11 pagesFimmu 13 848267Roii SianturiNo ratings yet
- Malaria in Humans Results From Infection by Any of Five Species of PlasmodiumDocument13 pagesMalaria in Humans Results From Infection by Any of Five Species of PlasmodiumDebabrata Ghosh DastidarNo ratings yet
- Transgenic Mosquitoes and The Fight AgainstDocument11 pagesTransgenic Mosquitoes and The Fight Againstlilibeth paola duran plataNo ratings yet
- IgG AntibodyDocument13 pagesIgG AntibodyerikaNo ratings yet
- Proposal EditedDocument7 pagesProposal EditedNtang MayaahNo ratings yet
- JurnalDocument19 pagesJurnalMOCHILNo ratings yet
- BEERNTSEN Et Al 2000Document23 pagesBEERNTSEN Et Al 2000Ana Beatriz Vasquez RodriguezNo ratings yet
- JDVAR 02 00037ooiDocument7 pagesJDVAR 02 00037ooiCrowdyMarineNo ratings yet
- Microevolution of Aedes Aegypti: Caroline Louise, Paloma Oliveira Vidal, Lincoln SuesdekDocument16 pagesMicroevolution of Aedes Aegypti: Caroline Louise, Paloma Oliveira Vidal, Lincoln SuesdekRocío Díaz GonzálezNo ratings yet
- Plasmodium Falciparum Gametocyte Dynamics in Areas of Different Malaria EndemicityDocument22 pagesPlasmodium Falciparum Gametocyte Dynamics in Areas of Different Malaria EndemicityAlemayehu Letebo AlbejoNo ratings yet
- Spatial Clustering of Fourteen Tick Species Across Districts of ZimbabweDocument9 pagesSpatial Clustering of Fourteen Tick Species Across Districts of ZimbabweMatthew ForestNo ratings yet
- Dengue Exposure and Wolbachia Wmel Strain Affects The Fertility of Quiescent Eggs of Aedes AegyptiDocument17 pagesDengue Exposure and Wolbachia Wmel Strain Affects The Fertility of Quiescent Eggs of Aedes AegyptiDANIEL ORTAÑEZNo ratings yet
- Genetic Approaches in Aedes Aegypti For Control of Dengue: An OverviewDocument12 pagesGenetic Approaches in Aedes Aegypti For Control of Dengue: An OverviewhechunNo ratings yet
- Lit - Jumbam - 2020 - BMC PH - KAP of Malaria Interventions in Rural ZambiaDocument15 pagesLit - Jumbam - 2020 - BMC PH - KAP of Malaria Interventions in Rural ZambianomdeplumNo ratings yet
- Research Progress On Surface Antigen 1 (SAG1) of Toxoplasma GondiiDocument9 pagesResearch Progress On Surface Antigen 1 (SAG1) of Toxoplasma Gondiiمحمود فضيلي ادم ابوالحسنNo ratings yet
- Jof 07 00844 v2Document11 pagesJof 07 00844 v2Sonia HernándezNo ratings yet
- Clase 1. Evolución de PatogenosDocument30 pagesClase 1. Evolución de PatogenosClaudia Alicia Herrera OlivasNo ratings yet
- 2022 Certad - CryptosopDocument10 pages2022 Certad - Cryptosopalexandraiuliana.aaNo ratings yet
- Review On Nanomaterials and Nano Scaled Systems For Topical and Systemic Delivery of Antifungal DrugsDocument23 pagesReview On Nanomaterials and Nano Scaled Systems For Topical and Systemic Delivery of Antifungal DrugsAnggun NurhidayahNo ratings yet
- The Genetics of Covid 19Document16 pagesThe Genetics of Covid 19Allan GuillermoNo ratings yet
- Review Article Sars Cov-2 and Other Reverse Zoonoses: Understanding The Human Animal Interface For Devising Control StrategiesDocument10 pagesReview Article Sars Cov-2 and Other Reverse Zoonoses: Understanding The Human Animal Interface For Devising Control Strategiescgoswami700No ratings yet
- Running Head: MALARIA 1Document7 pagesRunning Head: MALARIA 1klm klmNo ratings yet
- s41564-020-0690-4 SARS-CoV-2Document2 pagess41564-020-0690-4 SARS-CoV-2Juan Carlos GutiérrezNo ratings yet
- Biocontrol of Dengue-Draft CDocument3 pagesBiocontrol of Dengue-Draft Capi-685744638No ratings yet
- Al Amin 2020 PV 13 622Document15 pagesAl Amin 2020 PV 13 622poonamNo ratings yet
- Literatur WolbachiaDocument19 pagesLiteratur Wolbachialidya sianturiNo ratings yet
- s12936 023 04815 9Document12 pagess12936 023 04815 9Hosanna AGBANGLANo ratings yet
- 1Document13 pages1yowan wandikboNo ratings yet
- Ascaris y AscaridiasisDocument6 pagesAscaris y Ascaridiasisemmanueltobon0621No ratings yet
- Periodontal Infectogenomics: ReviewDocument6 pagesPeriodontal Infectogenomics: ReviewNadya PurwantyNo ratings yet
- Distintos Blancos Contra TicksDocument16 pagesDistintos Blancos Contra TicksSebastian MilesNo ratings yet
- Parasite 230005Document3 pagesParasite 230005caue1234No ratings yet
- SS BiochemDocument24 pagesSS BiochemQT네코No ratings yet
- Interactions Between Parasites and Human MicrobiotDocument6 pagesInteractions Between Parasites and Human MicrobiotGregorio De Las CasasNo ratings yet
- 2015, Aspergillus Fumigatus Mycovirus Causes Mild Hypervirulent Effect On Pathogenicity When Tested On Galleria MellonellaDocument7 pages2015, Aspergillus Fumigatus Mycovirus Causes Mild Hypervirulent Effect On Pathogenicity When Tested On Galleria MellonellaGorae DollNo ratings yet
- Plants 12 00793 v2Document4 pagesPlants 12 00793 v2Kath MadridNo ratings yet
- Host Specificity, Pathogen Exposure, and Superinfections Impact TheDocument14 pagesHost Specificity, Pathogen Exposure, and Superinfections Impact TheSarai MontielNo ratings yet
- Insect ImmunologyFrom EverandInsect ImmunologyNancy E. BeckageNo ratings yet
- 2020-Srikaya Seed Granule Different TemperatureDocument7 pages2020-Srikaya Seed Granule Different TemperatureDwiNo ratings yet
- A Transcriptomic Survey of TheDocument20 pagesA Transcriptomic Survey of TheDwiNo ratings yet
- 2014-Moringa Oleifera Trypsin Inh Dan Kill Bacteria MidgutDocument7 pages2014-Moringa Oleifera Trypsin Inh Dan Kill Bacteria MidgutDwiNo ratings yet
- 2014-Maybe Used-Ocotea CymbarumDocument10 pages2014-Maybe Used-Ocotea CymbarumDwiNo ratings yet
- 2017-Justicia Adhatoda MidgutDocument11 pages2017-Justicia Adhatoda MidgutDwiNo ratings yet
- Action of Natural PhytosanitarDocument10 pagesAction of Natural PhytosanitarDwiNo ratings yet
- A Transgenic Approach For ContDocument8 pagesA Transgenic Approach For ContDwiNo ratings yet
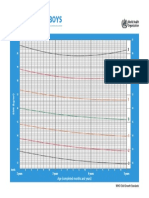
- CHT Bfa Boys Z 2 5Document1 pageCHT Bfa Boys Z 2 5DwiNo ratings yet
- A Toxin-Binding Alkaline PhospDocument20 pagesA Toxin-Binding Alkaline PhospDwiNo ratings yet
- A Transformed Bacterium ExpresDocument16 pagesA Transformed Bacterium ExpresDwiNo ratings yet
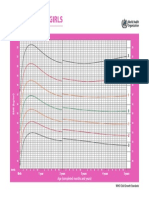
- CHT Bfa Girls Z 0 5Document1 pageCHT Bfa Girls Z 0 5DwiNo ratings yet
- Attachment 1706 631288cdc271dDocument5 pagesAttachment 1706 631288cdc271dDwiNo ratings yet
- CHT Bfa Boys Z 0 5Document1 pageCHT Bfa Boys Z 0 5DwiNo ratings yet
- 2012-Eichornia Crasiper-CulexDocument7 pages2012-Eichornia Crasiper-CulexDwiNo ratings yet
- Attachment 1706 631288cdc2e9fDocument4 pagesAttachment 1706 631288cdc2e9fDwiNo ratings yet
- 2012-CYP450 On EarthDocument12 pages2012-CYP450 On EarthDwiNo ratings yet
- 2012-Garlic-Allium Sativum EndotoxinDocument11 pages2012-Garlic-Allium Sativum EndotoxinDwiNo ratings yet
- 2012-Crimson Speckled MothDocument10 pages2012-Crimson Speckled MothDwiNo ratings yet
- 2012-Fungitoxic InsecticidalDocument18 pages2012-Fungitoxic InsecticidalDwiNo ratings yet
- 2012 Citrus Sinensis Musca DomesticaDocument8 pages2012 Citrus Sinensis Musca DomesticaDwiNo ratings yet
- 2012-Castor Bean Leaf-Spodoptera LituraDocument7 pages2012-Castor Bean Leaf-Spodoptera LituraDwiNo ratings yet
- Aedes Aegypti Bacillus ThuringiensisDocument9 pagesAedes Aegypti Bacillus ThuringiensisDwiNo ratings yet
- 2012-Bti SV2, Serratia Nematodipila SV6Document7 pages2012-Bti SV2, Serratia Nematodipila SV6DwiNo ratings yet
- 2012-Bti Residual TMOFDocument5 pages2012-Bti Residual TMOFDwiNo ratings yet
- 2012-Calatropis GiganteaDocument14 pages2012-Calatropis GiganteaDwiNo ratings yet
- 2012-Bti-Applied and Environmental MicrobiologyDocument6 pages2012-Bti-Applied and Environmental MicrobiologyDwiNo ratings yet
- The Genetic Basis of Dental Anomalies and Its Relation To OrthodonticsDocument5 pagesThe Genetic Basis of Dental Anomalies and Its Relation To Orthodonticsyuniarita eka putriNo ratings yet
- Midterm Topics Hema 1 LecDocument37 pagesMidterm Topics Hema 1 Lec4jzbxz64kqNo ratings yet
- Bacteria ProfileDocument18 pagesBacteria ProfileNOR-FATIMAH BARATNo ratings yet
- Troponin IDocument4 pagesTroponin IpetertrungNo ratings yet
- Bacterial Culture Guide ATCCDocument28 pagesBacterial Culture Guide ATCCAngel MurilloNo ratings yet
- Malaria Dengue Chikungunya Leptospirosis Hepatitis B Typhoid AgentDocument4 pagesMalaria Dengue Chikungunya Leptospirosis Hepatitis B Typhoid Agent70123No ratings yet
- Bacterial Disease AnalysisDocument63 pagesBacterial Disease AnalysisAshutoshkumar jhaNo ratings yet
- Salivary Gland Dysfunction: Etiology, Epidemiology, Clinical Manifestations, Diagnosis, and TreatmentDocument20 pagesSalivary Gland Dysfunction: Etiology, Epidemiology, Clinical Manifestations, Diagnosis, and TreatmentMonaNo ratings yet
- Protein SynthesisDocument14 pagesProtein SynthesisOginda MokoroNo ratings yet
- Atkins EnzimasDocument5 pagesAtkins EnzimasConstanza Espinoza LaraNo ratings yet
- 6 Hos CarcinogenesisDocument90 pages6 Hos CarcinogenesisalgutNo ratings yet
- Rett Syndrome A Neurological Disorder With MetabolDocument17 pagesRett Syndrome A Neurological Disorder With MetabolTiara PuspaNo ratings yet
- Answers To Multiple Choice Questions Test 2Document3 pagesAnswers To Multiple Choice Questions Test 2Nadeem IqbalNo ratings yet
- BrucellosisDocument65 pagesBrucellosisKotha Chaitanya AbhiramNo ratings yet
- Mulberry Genomics Progress and ProspectDocument21 pagesMulberry Genomics Progress and ProspectRay MrinalNo ratings yet
- Latoja, John Paul S. (50-Item Multiple Choice Test)Document12 pagesLatoja, John Paul S. (50-Item Multiple Choice Test)John Paul Sarmiento LatojaNo ratings yet
- BY, DR.M.L Kulkarni DR - Gayathri.K Davanagere. Reviewed by DR - Subramanya.N.K BangaloreDocument34 pagesBY, DR.M.L Kulkarni DR - Gayathri.K Davanagere. Reviewed by DR - Subramanya.N.K Bangaloregarlic100No ratings yet
- DermatologyDocument120 pagesDermatologyMustafa Thaier Al-KaraghouliNo ratings yet
- MIC Test Strip: Method PrincipleDocument8 pagesMIC Test Strip: Method PrincipleRuro LaudaNo ratings yet
- MARCH - 2023wDocument25 pagesMARCH - 2023wcariappa canvaNo ratings yet
- UTI TreatmentDocument3 pagesUTI TreatmentMaria RezitadinaNo ratings yet
- Malaria Drug Resistance in CameroonDocument93 pagesMalaria Drug Resistance in CameroonAnn TillNo ratings yet
- Amgen Inc. v. F. Hoffmann-LaRoche LTD Et Al - Document No. 1346Document12 pagesAmgen Inc. v. F. Hoffmann-LaRoche LTD Et Al - Document No. 1346Justia.comNo ratings yet
- Odontoblasts: The Cells Forming and Maintaining DentineDocument7 pagesOdontoblasts: The Cells Forming and Maintaining DentineJoniMukaNo ratings yet
- Product Catalogue: Keep Life FlowingDocument12 pagesProduct Catalogue: Keep Life FlowingTheencyclopediaNo ratings yet
- Chlorine Dioxide RoleDocument13 pagesChlorine Dioxide RolehanaNo ratings yet