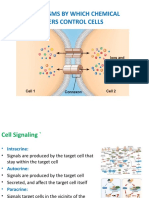
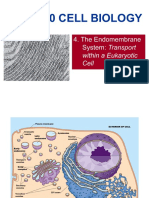
You might also like
- Biol2120 Cell Biology: Signal Transduction Mechanisms: II. Messengers and ReceptorsDocument58 pagesBiol2120 Cell Biology: Signal Transduction Mechanisms: II. Messengers and ReceptorsHui Ka HoNo ratings yet
- Ch5 Cell SignallingDocument50 pagesCh5 Cell SignallingShatha KhzaimiahNo ratings yet
- Signal TransductionDocument30 pagesSignal Transductionpalak100% (1)
- Signal Transduction Mechanisms and Cellular ResponsesDocument39 pagesSignal Transduction Mechanisms and Cellular ResponsesBellony Sanders100% (1)
- Cell Signaling I: Signal Transduction & Short-term Cellular ResponsesDocument40 pagesCell Signaling I: Signal Transduction & Short-term Cellular ResponsesRNo ratings yet
- Lecture 5Document26 pagesLecture 5BlaNo ratings yet
- Cell SignalingDocument45 pagesCell SignalingNand PrakashNo ratings yet
- Cell SignalingDocument53 pagesCell SignalingKaushiki KalravNo ratings yet
- Communicating Cells: How Cells Send and Receive SignalsDocument46 pagesCommunicating Cells: How Cells Send and Receive SignalsFachriza EffendiNo ratings yet
- Module 5Document108 pagesModule 5alshalabinezardonia96No ratings yet
- CB CS07 CellCommunicationDocument1 pageCB CS07 CellCommunicationstcgtdNo ratings yet
- G-Protein Coupled ReceptorsDocument8 pagesG-Protein Coupled ReceptorsHyunji KimNo ratings yet
- ch9 SlidesDocument18 pagesch9 Slidesapi-197124028No ratings yet
- Drug Receptor Bonding Signal Transduction 05mars2012 10Document34 pagesDrug Receptor Bonding Signal Transduction 05mars2012 10Yatish KarjekarNo ratings yet
- Cell Communication SignalsDocument27 pagesCell Communication Signalsbostonsox49No ratings yet
- Intracellular SignallingDocument45 pagesIntracellular Signallingsana iqbalNo ratings yet
- 3,4-Receptors As Drug TargetsDocument166 pages3,4-Receptors As Drug TargetsyudenfranciscoNo ratings yet
- Mechanism of Hormone Action: DR Jehad Al-ShuneigatDocument47 pagesMechanism of Hormone Action: DR Jehad Al-Shuneigatyousef sarairehNo ratings yet
- Membrane Receptors and Their PhysiologyDocument32 pagesMembrane Receptors and Their Physiologygee kodexNo ratings yet
- ReceptorsDocument31 pagesReceptorsBareera NudratNo ratings yet
- Cell SignalingDocument75 pagesCell SignalingjhanvisNo ratings yet
- Unit 4 PPT 1 Topics 4.1 - 4.4 Student NotesDocument30 pagesUnit 4 PPT 1 Topics 4.1 - 4.4 Student Notesavalon.protter08No ratings yet
- Cell Communication CH 11Document43 pagesCell Communication CH 11suryani yaniNo ratings yet
- Cell Signaling: Signaling Molecules and Their ReceptorsDocument49 pagesCell Signaling: Signaling Molecules and Their ReceptorsDeepanshu rawatNo ratings yet
- Plasma Membrane ReceptorDocument10 pagesPlasma Membrane ReceptorAkhilesh BhuraNo ratings yet
- F22 MCB 2050 Lecture 10 - Signal TransductionDocument28 pagesF22 MCB 2050 Lecture 10 - Signal TransductionNO VIDEOSNo ratings yet
- 2-Hormonal Signal Transduction 2 2Document33 pages2-Hormonal Signal Transduction 2 2Mohammed Mansour AbdullahNo ratings yet
- Chapter 7: Cell Communication and Signaling 1: Dr. Ivonne BejaranoDocument29 pagesChapter 7: Cell Communication and Signaling 1: Dr. Ivonne BejaranossaNo ratings yet
- Pharmacodynamics of Drug ReceptorsDocument11 pagesPharmacodynamics of Drug ReceptorsMandyNo ratings yet
- Receptors & Signal TransductionDocument10 pagesReceptors & Signal TransductionArslan SaeedNo ratings yet
- ReceptorsDocument16 pagesReceptorsM HarisNo ratings yet
- BiosignallingDocument16 pagesBiosignallingAsmaa HatemNo ratings yet
- Cell To Cell CommunicationDocument29 pagesCell To Cell CommunicationSheerin SulthanaNo ratings yet
- Cell Communication SignalsDocument41 pagesCell Communication SignalskomorebuNo ratings yet
- BABS2202 Immunology and Cell Signalling Study GuideDocument5 pagesBABS2202 Immunology and Cell Signalling Study GuideDimitri KandilasNo ratings yet
- Medicinal Chemistry:-Receptors:-: LigandDocument9 pagesMedicinal Chemistry:-Receptors:-: LigandgalooangelNo ratings yet
- Cell and Molecular Biology: Gerald KarpDocument53 pagesCell and Molecular Biology: Gerald Karppat ongNo ratings yet
- Neurotransmitter Cell CommunicationDocument25 pagesNeurotransmitter Cell CommunicationOtnil DNo ratings yet
- Lect. 10Document27 pagesLect. 10أحمدشريفNo ratings yet
- Cell Signaling Signal TransductionDocument7 pagesCell Signaling Signal TransductionArslan AliNo ratings yet
- Receptor Mechanisms: - Receptor Are Proteins With High Affinity To ADocument45 pagesReceptor Mechanisms: - Receptor Are Proteins With High Affinity To AbevorsiNo ratings yet
- CP3 - Intracellular Signaling at The Cell MembraneDocument31 pagesCP3 - Intracellular Signaling at The Cell Membranedafabc50No ratings yet
- Molecular Mechanisms of Drug ActionDocument76 pagesMolecular Mechanisms of Drug ActionSunil100% (1)
- Mechanism of Hormone ActionDocument30 pagesMechanism of Hormone ActionWhite SnelliNo ratings yet
- Chapt09 Lecture 2019-SDocument45 pagesChapt09 Lecture 2019-SPaulNo ratings yet
- CH 11 PPT Cell Communication 1Document77 pagesCH 11 PPT Cell Communication 1api-270681964No ratings yet
- Receptors IntroductionDocument53 pagesReceptors IntroductionSunilNo ratings yet
- Mechanism of Hormone ActionDocument54 pagesMechanism of Hormone ActionDr. Tapan Kr. Dutta100% (1)
- Cell CommunicationDocument24 pagesCell Communicationanbuselvanselvan59No ratings yet
- Pharmacodynamics Effects and MechanismsDocument76 pagesPharmacodynamics Effects and Mechanismsperala vinaykumarNo ratings yet
- Receptor SuperfamiliesDocument27 pagesReceptor SuperfamiliesNAUSHEEN SULTANANo ratings yet
- Govt. Digvijay Autonomous P. G. College Raj-Nandgaon (C. G.)Document27 pagesGovt. Digvijay Autonomous P. G. College Raj-Nandgaon (C. G.)oliver senNo ratings yet
- Lecture 4Document32 pagesLecture 4Muhanad Shahin100% (1)
- Genetic variation in GPCR may impact diseases and drug responseDocument17 pagesGenetic variation in GPCR may impact diseases and drug responseSiddharth Shekhar SinghNo ratings yet
- Signal TransductionDocument13 pagesSignal Transductionakanksha kapoorNo ratings yet
- Pharmacodynamics: DR Narendra KumarDocument76 pagesPharmacodynamics: DR Narendra KumarsivaNo ratings yet
- 6 General SignallingDocument14 pages6 General SignallingBill John PedregosaNo ratings yet
- ReceptorsDocument44 pagesReceptorsAbiy AliyeNo ratings yet
- 2c-MOLECULAR ASPECTS OF DRUG ACTIONDocument66 pages2c-MOLECULAR ASPECTS OF DRUG ACTIONAlly AbdalahNo ratings yet
- Biol2120 Cell Biology: 8. Apoptosis: TheDocument28 pagesBiol2120 Cell Biology: 8. Apoptosis: TheHui Ka HoNo ratings yet
- BIOL2120 7 Cell CycleDocument44 pagesBIOL2120 7 Cell CycleHui Ka HoNo ratings yet
- Biol2120 Cell Biology: Structures For Maintaining Cell Shape and Internal OrganizationDocument49 pagesBiol2120 Cell Biology: Structures For Maintaining Cell Shape and Internal OrganizationHui Ka HoNo ratings yet
- BIOL2120 1 IntroductionDocument26 pagesBIOL2120 1 IntroductionHui Ka HoNo ratings yet
- Cancer Cells - Understanding the Biology of Cancer at the Cellular LevelDocument28 pagesCancer Cells - Understanding the Biology of Cancer at the Cellular LevelHui Ka HoNo ratings yet
- Signal Transduction: Messengers, Receptors and Cellular ResponsesDocument58 pagesSignal Transduction: Messengers, Receptors and Cellular ResponsesHui Ka HoNo ratings yet
- BIOL2120 7 Cell CycleDocument44 pagesBIOL2120 7 Cell CycleHui Ka HoNo ratings yet
- BIOL2120 1 IntroductionDocument26 pagesBIOL2120 1 IntroductionHui Ka HoNo ratings yet
- BIOL2120 3 Transport Across MembranesDocument60 pagesBIOL2120 3 Transport Across MembranesHui Ka HoNo ratings yet
- BIOL2120 4 Endomembrane SystemDocument47 pagesBIOL2120 4 Endomembrane SystemHui Ka HoNo ratings yet
- BIOL2120 4 Endomembrane SystemDocument47 pagesBIOL2120 4 Endomembrane SystemHui Ka HoNo ratings yet
- Biol2120 Cell Biology: Membranes: Their Structure, Function, and Chemistry 2. MembranesDocument49 pagesBiol2120 Cell Biology: Membranes: Their Structure, Function, and Chemistry 2. MembranesHui Ka HoNo ratings yet
- Biol2120 Cell Biology: 8. Apoptosis: TheDocument28 pagesBiol2120 Cell Biology: 8. Apoptosis: TheHui Ka HoNo ratings yet
- Biol2120 Cell Biology: 8. Apoptosis: TheDocument28 pagesBiol2120 Cell Biology: 8. Apoptosis: TheHui Ka HoNo ratings yet
- Cytoskeleton Structures for Cell Shape and OrganizationDocument49 pagesCytoskeleton Structures for Cell Shape and OrganizationHui Ka HoNo ratings yet
- BIOL2120 3 Transport Across MembranesDocument60 pagesBIOL2120 3 Transport Across MembranesHui Ka HoNo ratings yet
- Cancer Cells - Understanding the Biology of Cancer at the Cellular LevelDocument28 pagesCancer Cells - Understanding the Biology of Cancer at the Cellular LevelHui Ka HoNo ratings yet
- Cancer Cells - Understanding the Biology of Cancer at the Cellular LevelDocument28 pagesCancer Cells - Understanding the Biology of Cancer at the Cellular LevelHui Ka HoNo ratings yet
- BIOL2120 1 IntroductionDocument26 pagesBIOL2120 1 IntroductionHui Ka HoNo ratings yet
- Biol2120 Cell Biology: Structures For Maintaining Cell Shape and Internal OrganizationDocument49 pagesBiol2120 Cell Biology: Structures For Maintaining Cell Shape and Internal OrganizationHui Ka HoNo ratings yet
- Biol2120 Cell Biology: Membranes: Their Structure, Function, and Chemistry 2. MembranesDocument49 pagesBiol2120 Cell Biology: Membranes: Their Structure, Function, and Chemistry 2. MembranesHui Ka HoNo ratings yet
- BIOL2120 3 Transport Across MembranesDocument60 pagesBIOL2120 3 Transport Across MembranesHui Ka HoNo ratings yet
- Signal Transduction: Messengers, Receptors and Cellular ResponsesDocument58 pagesSignal Transduction: Messengers, Receptors and Cellular ResponsesHui Ka HoNo ratings yet
- BIOL2120 4 Endomembrane SystemDocument47 pagesBIOL2120 4 Endomembrane SystemHui Ka HoNo ratings yet
- BIOL2120 7 Cell CycleDocument44 pagesBIOL2120 7 Cell CycleHui Ka HoNo ratings yet
- BIOL2120 1 IntroductionDocument26 pagesBIOL2120 1 IntroductionHui Ka HoNo ratings yet
- Cancer Cells - Understanding the Biology of Cancer at the Cellular LevelDocument28 pagesCancer Cells - Understanding the Biology of Cancer at the Cellular LevelHui Ka HoNo ratings yet
- Biol2120 Cell Biology: Membranes: Their Structure, Function, and Chemistry 2. MembranesDocument49 pagesBiol2120 Cell Biology: Membranes: Their Structure, Function, and Chemistry 2. MembranesHui Ka HoNo ratings yet
- BIO2120 Cell Death Programmed ApoptosisDocument28 pagesBIO2120 Cell Death Programmed ApoptosisHui Ka HoNo ratings yet
- Basic terms in biochemistry explainedDocument41 pagesBasic terms in biochemistry explainedkumarklNo ratings yet
- Biomolecules Chemistry AssignmentDocument28 pagesBiomolecules Chemistry AssignmentAnurag Singh Thakur67% (3)
- Cell Structure & Cell Cycle QuestionsDocument21 pagesCell Structure & Cell Cycle QuestionsTassie777No ratings yet
- BCHM 461 Exam #2 Problem SolutionsDocument5 pagesBCHM 461 Exam #2 Problem SolutionsSurajit BhattacharjeeNo ratings yet
- Bio MoleculeDocument14 pagesBio MoleculeMohit KaushikNo ratings yet
- Primer Designing For PCR: ColloquiumDocument3 pagesPrimer Designing For PCR: ColloquiumSakshi SangleNo ratings yet
- Biochem Lab Report 2Document8 pagesBiochem Lab Report 2Lih XuanNo ratings yet
- Devlin Biochemistry With Clinical CorrelationsDocument1,241 pagesDevlin Biochemistry With Clinical CorrelationsJorge100% (5)
- Lipid MetabolismDocument25 pagesLipid Metabolismബാദ്ഷാ ഖാൻNo ratings yet
- Food Technology XLDocument113 pagesFood Technology XLSubhankar MaityNo ratings yet
- A1.2 Nucleic AcidsDocument83 pagesA1.2 Nucleic AcidsGhenwa DimachkiNo ratings yet
- Molecular Biology QuestionsDocument6 pagesMolecular Biology QuestionsVinay Kumar100% (1)
- Fatty Acid Oxidation Part Two Illustration AtfDocument1 pageFatty Acid Oxidation Part Two Illustration AtfDivyaa VisvalingamNo ratings yet
- Protein AggregationDocument29 pagesProtein AggregationAndré PérezNo ratings yet
- 2.1.2 Biological Molecules ChecklistDocument2 pages2.1.2 Biological Molecules Checklistrashahocine07No ratings yet
- RNAProtein Synthesis SEDocument7 pagesRNAProtein Synthesis SEAmbrielle WhiteNo ratings yet
- Human Hair To Protien ThesiDocument53 pagesHuman Hair To Protien ThesiTANMAY DALVINo ratings yet
- 12-4 Gene Regulation and MutationDocument26 pages12-4 Gene Regulation and Mutationapi-342334216No ratings yet
- Biology 111 EXAM 2 Study GuideDocument15 pagesBiology 111 EXAM 2 Study GuidecuckoofourkokoNo ratings yet
- Fundamental Molecular Biology: Lisabeth A. AllisonDocument73 pagesFundamental Molecular Biology: Lisabeth A. AllisonJu noNo ratings yet
- FEBS Open Bio - 2023 - Hampel - CoCl2 Triggered Pseudohypoxic Stress Induces Proteasomal Degradation of SIRT4 ViaDocument13 pagesFEBS Open Bio - 2023 - Hampel - CoCl2 Triggered Pseudohypoxic Stress Induces Proteasomal Degradation of SIRT4 ViaMallik ArjunNo ratings yet
- Antibody Production and Purification Technical Handbook, Version 2Document41 pagesAntibody Production and Purification Technical Handbook, Version 2bh14561No ratings yet
- Essential Biochemistry - S PDFDocument586 pagesEssential Biochemistry - S PDFMariam MA100% (1)
- Protein Denaturation FactorsDocument3 pagesProtein Denaturation FactorsJemaica A. JagolinoNo ratings yet
- Proteins Practice QuizDocument5 pagesProteins Practice Quizafaflotfi_155696459No ratings yet
- Amino AcidsDocument6 pagesAmino AcidsJohannaNo ratings yet
- Hole's Human Anatomy and Physiology: Shier Butler LewisDocument12 pagesHole's Human Anatomy and Physiology: Shier Butler LewisTheophilus BaidooNo ratings yet
- AP Biology Dna SGDocument3 pagesAP Biology Dna SGFaisaludinNo ratings yet
- Energy Balance On Separator (Correction)Document4 pagesEnergy Balance On Separator (Correction)nabilaNo ratings yet
- Rubric For DNA and RNA ModelDocument2 pagesRubric For DNA and RNA ModelDrew MarasiganNo ratings yet