You might also like
- Human Chromosomes: An Illustrated Introduction to Human CytogeneticsFrom EverandHuman Chromosomes: An Illustrated Introduction to Human CytogeneticsRating: 5 out of 5 stars5/5 (1)
- Centrosome Inheritance After Fertilization and Nuclear Transfer in MammalsDocument14 pagesCentrosome Inheritance After Fertilization and Nuclear Transfer in MammalsBlahNo ratings yet
- Kinetochore Assembly and Function Through The Cell Cycle: ReviewDocument15 pagesKinetochore Assembly and Function Through The Cell Cycle: ReviewLê Khánh ToànNo ratings yet
- Centrosome DinamicsDocument9 pagesCentrosome DinamicsEugenia Alaniz FierroNo ratings yet
- Aurora & MitosisDocument11 pagesAurora & MitosisJubairNo ratings yet
- Aurora Kinases, Aneuploidy and Cancer: A Real LinkDocument10 pagesAurora Kinases, Aneuploidy and Cancer: A Real LinkalmaNo ratings yet
- Symmetry-Breaking of Animal CytokinesisDocument10 pagesSymmetry-Breaking of Animal CytokinesisLizbeth EspinosaNo ratings yet
- R2 FrontiersDocument18 pagesR2 FrontiersRK VermaNo ratings yet
- Cytoskeleton - 2012 - Farr - Cytokinesis in TrypanosomesDocument11 pagesCytoskeleton - 2012 - Farr - Cytokinesis in TrypanosomesDario DiosNo ratings yet
- Centrosome - Wikipedia, The Free EncyclopediaDocument6 pagesCentrosome - Wikipedia, The Free EncyclopediaShailendra YadavNo ratings yet
- Pbifvjeoiif Ki Ojo LKJ JKNJ NJ Noi Jo Joi Jio Jiu JDocument3 pagesPbifvjeoiif Ki Ojo LKJ JKNJ NJ Noi Jo Joi Jio Jiu JLaia Baus RevueltaNo ratings yet
- The Bacterial Cytoskeleton 1Document2 pagesThe Bacterial Cytoskeleton 1EdnaCharryNo ratings yet
- Nuclear Envelope Attachment Is Not Necessary For Telomere Function in Fission YeastDocument6 pagesNuclear Envelope Attachment Is Not Necessary For Telomere Function in Fission YeastLê Khánh ToànNo ratings yet
- About Aurora KinaseDocument6 pagesAbout Aurora KinaseJubairNo ratings yet
- Schenkel Et Al 2023 A Dedicated Cytoplasmic Container Collects Extrachromosomal Dna Away From The Mammalian NucleusDocument21 pagesSchenkel Et Al 2023 A Dedicated Cytoplasmic Container Collects Extrachromosomal Dna Away From The Mammalian NucleusGuilhermeNo ratings yet
- 1999-Identification and Function of The Centrosome CentromatrixDocument10 pages1999-Identification and Function of The Centrosome CentromatrixyicinenNo ratings yet
- The Chromosomal Passenger Complex (CPC) : From Easy Rider To The Godfather of MitosisDocument15 pagesThe Chromosomal Passenger Complex (CPC) : From Easy Rider To The Godfather of MitosisRK VermaNo ratings yet
- Cell division and the cell cycle explainedDocument15 pagesCell division and the cell cycle explainedadri baigorriNo ratings yet
- Centriol and CiliaDocument27 pagesCentriol and CiliahamdyyalyNo ratings yet
- Definition RevisedDocument7 pagesDefinition Revisedapi-406572841No ratings yet
- Ultrastructure Nucleus: BiologyDocument14 pagesUltrastructure Nucleus: BiologyAshu ShewaleNo ratings yet
- Chapter 9: The Cell Cycle and Cellular Reproduction 9.1 The Cell CycleDocument4 pagesChapter 9: The Cell Cycle and Cellular Reproduction 9.1 The Cell CycleJennifer Muñoz ValenciaNo ratings yet
- Cell division and the cell cycleDocument13 pagesCell division and the cell cycleNicholasNo ratings yet
- R2 Frontiers2017Document21 pagesR2 Frontiers2017RK VermaNo ratings yet
- Nihms 367187Document14 pagesNihms 367187Tiego FerreiraNo ratings yet
- Cell and Cell Division: FBS 74 Forest Genetics and Tree ImprovementDocument6 pagesCell and Cell Division: FBS 74 Forest Genetics and Tree ImprovementFaith Hope NavarraNo ratings yet
- 1.6 Cell DivisionDocument46 pages1.6 Cell DivisionJoanita Jasmine JeniferNo ratings yet
- BZ Lab 3Document11 pagesBZ Lab 3Alexa Jean D. HonrejasNo ratings yet
- Topic: Cytokinesis: Etymology and PronunciationDocument6 pagesTopic: Cytokinesis: Etymology and PronunciationVindhya ShankerNo ratings yet
- CH - 44 - Mitosis and Cytokinesis PDFDocument24 pagesCH - 44 - Mitosis and Cytokinesis PDFerichaasNo ratings yet
- Jurnal Genetikaenglish 2Document9 pagesJurnal Genetikaenglish 2Leonard SitumorangNo ratings yet
- Cell Division: Understanding Mitosis and Its PhasesDocument42 pagesCell Division: Understanding Mitosis and Its PhasesReeya WhabiNo ratings yet
- Bab 1 2 Biomembran PDFDocument84 pagesBab 1 2 Biomembran PDFMuh JufriNo ratings yet
- JCB 202209062Document21 pagesJCB 202209062Aleksay NesmashnyyNo ratings yet
- Baghdad College of Medical Science (Human Biology)Document9 pagesBaghdad College of Medical Science (Human Biology)علي صلاح مهديNo ratings yet
- Sitokinesis Pada Proses Pembelahan Sel MamaliaDocument6 pagesSitokinesis Pada Proses Pembelahan Sel MamaliaAnonymous AkCt1GONo ratings yet
- Lecture 3 - Chapter 8-Cytoskeleton ADocument75 pagesLecture 3 - Chapter 8-Cytoskeleton AKw Chan33% (3)
- ISSCR WriteupDocument3 pagesISSCR Writeupachuthasaran@11No ratings yet
- An Aurora Kinase Inhibitor, AMG900, Inhibits Glioblastoma Cell Proliferation by Disrupting Mitotic ProgressionDocument15 pagesAn Aurora Kinase Inhibitor, AMG900, Inhibits Glioblastoma Cell Proliferation by Disrupting Mitotic ProgressionJubairNo ratings yet
- Self-Organized Nuclear Positioning Synchronizes The Cell Cycle in Drosophila EmbryosDocument35 pagesSelf-Organized Nuclear Positioning Synchronizes The Cell Cycle in Drosophila EmbryosエリタノアンソネーNo ratings yet
- The Actin Cortex at A GlanceDocument9 pagesThe Actin Cortex at A GlanceArkayan LahaNo ratings yet
- Vale Cell 2003 Review Molecular Motor Toolbox For Intracellular TransportDocument14 pagesVale Cell 2003 Review Molecular Motor Toolbox For Intracellular TransportSagarNo ratings yet
- 0000 2005 Nature NSC Cell and NeurogenesisDocument12 pages0000 2005 Nature NSC Cell and NeurogenesisEsteban Daniel Peluffo ArgónNo ratings yet
- Cho2016 Article RolesOfCross-MembraneTransportDocument13 pagesCho2016 Article RolesOfCross-MembraneTransportTaurus PearsonNo ratings yet
- BioK DP Notes 1.6Document13 pagesBioK DP Notes 1.6Basira GasimliNo ratings yet
- Cells 1367329Document19 pagesCells 1367329alvand gharibNo ratings yet
- Understanding the Dynamic CytoskeletonDocument66 pagesUnderstanding the Dynamic CytoskeletonChristian Santiago100% (1)
- Article32 PDFDocument4 pagesArticle32 PDFdupuytrenNo ratings yet
- Genetics Lec - Quiz 1 HandoutsDocument7 pagesGenetics Lec - Quiz 1 Handoutsvada_soNo ratings yet
- Act04 Cell Division - Mitosis DiscussionDocument3 pagesAct04 Cell Division - Mitosis DiscussionPaula Nicole AlanoNo ratings yet
- Lecture III - Semi-FinalsDocument16 pagesLecture III - Semi-FinalsENGETS IT7No ratings yet
- Cell Nucleus, Mitochondrium, Peroxysome: Viktoria Vereczki, M.D., PH.DDocument47 pagesCell Nucleus, Mitochondrium, Peroxysome: Viktoria Vereczki, M.D., PH.D20SMB24 - ShafanaNo ratings yet
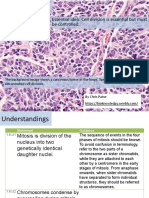
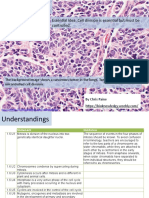
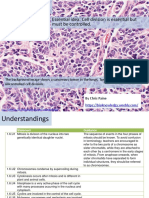
- 1.6 Cell Division: Essential Idea: Cell Division Is Essential But Must Be ControlledDocument33 pages1.6 Cell Division: Essential Idea: Cell Division Is Essential But Must Be ControlledAnonymous bOMSH4wDgeNo ratings yet
- Alternatives To Binary Fission in BacteriaDocument11 pagesAlternatives To Binary Fission in BacteriaCLAUDIA SICHELNo ratings yet
- Cell Cycle and Division NotesDocument5 pagesCell Cycle and Division NotesMicaela DavisNo ratings yet
- 1.6 Cell DivisionDocument30 pages1.6 Cell DivisionAnnabeth ParkerNo ratings yet
- Chapter 06Document39 pagesChapter 06NnleinomNo ratings yet
- Lab 9Document40 pagesLab 9bob0% (1)
- J.A. Tuszynski, J.A. Brown, E. Crawford, E.J. Carpenter, M.L.A. Nip, J.M. Dixon and M.V. Sataric: Molecular Dynamics Simulations of Tubulin Structure and Calculations of Electrostatic Properties of MicrotubulesDocument16 pagesJ.A. Tuszynski, J.A. Brown, E. Crawford, E.J. Carpenter, M.L.A. Nip, J.M. Dixon and M.V. Sataric: Molecular Dynamics Simulations of Tubulin Structure and Calculations of Electrostatic Properties of MicrotubulesUloffNo ratings yet
- 1 The Cell-Cycle Control MorganDocument2 pages1 The Cell-Cycle Control MorganalmaNo ratings yet
- 1 Cdks and Cyclins G1-Differentiation-Embryonic-Hematopoietic 010Document8 pages1 Cdks and Cyclins G1-Differentiation-Embryonic-Hematopoietic 010alma ramirezNo ratings yet
- Apoptosis-Induced Compensatory Proliferation08Document7 pagesApoptosis-Induced Compensatory Proliferation08almaNo ratings yet
- How Do BCL-2 Proteins Induce Mitochondrial-Membrane Permeabiliza08Document8 pagesHow Do BCL-2 Proteins Induce Mitochondrial-Membrane Permeabiliza08almaNo ratings yet
- Aurora Meiosis and Mitosis 03Document15 pagesAurora Meiosis and Mitosis 03almaNo ratings yet
- Atlas HamsterDocument75 pagesAtlas Hamsteralma100% (1)
- Desarrollo de Oocitos en RhesusDocument8 pagesDesarrollo de Oocitos en RhesusalmaNo ratings yet
- 101Document12 pages101ilbichNo ratings yet
- Meghan Markle In-N-Out Burger Photo Deepens Mystery of Why She's Not With Prince Harry - News - Com.au - Australia's Leading News SiteDocument10 pagesMeghan Markle In-N-Out Burger Photo Deepens Mystery of Why She's Not With Prince Harry - News - Com.au - Australia's Leading News SiteTia XNo ratings yet
- A. Fill The Picture Below With Correct Greetings. (6 Marks) Good Morning Hello Good Evening Good Afternoon Good Night GoodbyeDocument8 pagesA. Fill The Picture Below With Correct Greetings. (6 Marks) Good Morning Hello Good Evening Good Afternoon Good Night GoodbyeBandan UnyatNo ratings yet
- Report-Gaps in Policies and Laws That Perpertuate GBV in TanzaniaDocument32 pagesReport-Gaps in Policies and Laws That Perpertuate GBV in TanzaniaPaschal KunambiNo ratings yet
- Relationships: Latin Key WordsDocument2 pagesRelationships: Latin Key WordsIanua ViteNo ratings yet
- Dental and Vision Enrollment FormDocument3 pagesDental and Vision Enrollment Formsatnam_25No ratings yet
- Mary Queen of Scots Events 1542-1566Document9 pagesMary Queen of Scots Events 1542-1566Arif NadafNo ratings yet
- Scottish Notes and Queries: Is Given Only in Conventional Upper/LowercaseDocument4 pagesScottish Notes and Queries: Is Given Only in Conventional Upper/LowercaseSommerledNo ratings yet
- Unit 1 I. Fill in With The Appropriate WordDocument2 pagesUnit 1 I. Fill in With The Appropriate WordAndreea RaduNo ratings yet
- Primary Healthcare Centre ServicesDocument6 pagesPrimary Healthcare Centre ServicesAnkitChauhanNo ratings yet
- Digital Booklet - Britney Spears - Gimme More (Digital 45)Document2 pagesDigital Booklet - Britney Spears - Gimme More (Digital 45)Gâu Ehehe50% (2)
- Harry Potter ProjectDocument21 pagesHarry Potter ProjectGabrielaMoriliaNo ratings yet
- Sample Template For ThesisDocument8 pagesSample Template For ThesisJordan CruzNo ratings yet
- SCRUM MASTER - Master of Stances 1Document2 pagesSCRUM MASTER - Master of Stances 1shikha kathuriaNo ratings yet
- Sho Ogawa's Thesis On Pinku Eiga FilmsDocument88 pagesSho Ogawa's Thesis On Pinku Eiga FilmsbrozacaNo ratings yet
- ACTS 365 (Demographic Statistics)Document117 pagesACTS 365 (Demographic Statistics)Prince KayNo ratings yet
- Edac225 Assignment 1c - Case StudyDocument7 pagesEdac225 Assignment 1c - Case Studyapi-319193686No ratings yet
- Arm Wrestling With My Father PDFDocument4 pagesArm Wrestling With My Father PDFIjyel VillaNo ratings yet
- Anaya Vs PalaroanDocument2 pagesAnaya Vs PalaroanTippy Dos SantosNo ratings yet
- V-7 G.R. No. 150611 Saguid Vs CADocument7 pagesV-7 G.R. No. 150611 Saguid Vs CAIter MercatabantNo ratings yet
- Noveras V Noveras GR 188289 08202014Document9 pagesNoveras V Noveras GR 188289 08202014Dorothy Joy LucmayonNo ratings yet
- Habeas CorpusDocument11 pagesHabeas CorpusAnathea CadagatNo ratings yet
- Municipal Ordinance No. 04-2012Document6 pagesMunicipal Ordinance No. 04-2012JayPardinianNuyda100% (1)
- SB 133Document5 pagesSB 133Lindsey BasyeNo ratings yet
- Legal Separation CasesDocument52 pagesLegal Separation CasesCrnc NavidadNo ratings yet
- Ravished by The Wolf Alpha Male Succub Alexander AshleeDocument162 pagesRavished by The Wolf Alpha Male Succub Alexander AshleeKathy030878% (9)
- Right of Children (Maintenance, Inheritance, Right of Illegitimate Children, Adoption Etc.) :prepared By: H. S. MuliaDocument60 pagesRight of Children (Maintenance, Inheritance, Right of Illegitimate Children, Adoption Etc.) :prepared By: H. S. MuliaAman jainNo ratings yet
- Civ DigestDocument2 pagesCiv DigestSusan Sabilala MangallenoNo ratings yet
- OtherDocument5 pagesOtherA.P. MadarangNo ratings yet
- University of The Philippines College of LawDocument5 pagesUniversity of The Philippines College of Lawrm2803No ratings yet
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (378)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Inside of a Dog: What Dogs See, Smell, and KnowFrom EverandInside of a Dog: What Dogs See, Smell, and KnowRating: 4 out of 5 stars4/5 (390)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- Why We Sleep: Unlocking the Power of Sleep and DreamsFrom EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsRating: 4.5 out of 5 stars4.5/5 (2083)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (3)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- The Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsFrom EverandThe Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsNo ratings yet
- The Invention of Tomorrow: A Natural History of ForesightFrom EverandThe Invention of Tomorrow: A Natural History of ForesightRating: 4.5 out of 5 stars4.5/5 (5)