You might also like
- Centrosome Inheritance After Fertilization and Nuclear Transfer in MammalsDocument14 pagesCentrosome Inheritance After Fertilization and Nuclear Transfer in MammalsBlahNo ratings yet
- Cellular Differentiation: Mammalian Cell TypesDocument7 pagesCellular Differentiation: Mammalian Cell TypesMuhammad ZairulfikriNo ratings yet
- Mitosis Is The Process by Which A: Aspergillus Nidulans Saccharomyces CerevisiaeDocument7 pagesMitosis Is The Process by Which A: Aspergillus Nidulans Saccharomyces CerevisiaeLeo Paolo LuceroNo ratings yet
- Mitosis: Not To Be Confused With, ,, orDocument6 pagesMitosis: Not To Be Confused With, ,, orWacek4444No ratings yet
- Definition RevisedDocument7 pagesDefinition Revisedapi-406572841No ratings yet
- Cellular Differentiation: Mammalian Cell TypesDocument7 pagesCellular Differentiation: Mammalian Cell TypesMuhammad ZairulfikriNo ratings yet
- MitosisDocument3 pagesMitosisVinodh RavindaranNo ratings yet
- 1.6 Cell DivisionDocument46 pages1.6 Cell DivisionJoanita Jasmine JeniferNo ratings yet
- 0 BooksDocument20 pages0 Bookslaraib zahidNo ratings yet
- Mitosis - WikipediaDocument107 pagesMitosis - WikipediaBashiir NuurNo ratings yet
- 2.5 - Cell DivisionDocument5 pages2.5 - Cell DivisionAlvin JovanNo ratings yet
- Phases of The Cell CycleDocument9 pagesPhases of The Cell CycleZia PhotostateNo ratings yet
- Final Examination-BiologyDocument15 pagesFinal Examination-BiologyAlliah MendozaNo ratings yet
- Cell DivisionDocument17 pagesCell DivisionGenevieve GanubNo ratings yet
- MitosisDocument15 pagesMitosisJorge MurieteNo ratings yet
- CH8 Cell DivisionsDocument69 pagesCH8 Cell DivisionsAhmed MohamadNo ratings yet
- Biology ProjectDocument27 pagesBiology ProjectRajatNo ratings yet
- Chapter Ten:: Cell Cycle and Cell DivisionDocument12 pagesChapter Ten:: Cell Cycle and Cell DivisionShivani Shree SundaramoorthyNo ratings yet
- Zoology Lecture 5Document7 pagesZoology Lecture 5Stephen OcheaNo ratings yet
- Lab 9Document40 pagesLab 9bob0% (1)
- Mitosis - WikipediaDocument39 pagesMitosis - WikipediaMuhammad Azhar MagsiNo ratings yet
- 1.6 Cell DivisionDocument30 pages1.6 Cell DivisionAnnabeth ParkerNo ratings yet
- Chapter 21Document20 pagesChapter 21Uzma AdnanNo ratings yet
- Centrosome DinamicsDocument9 pagesCentrosome DinamicsEugenia Alaniz FierroNo ratings yet
- רזי 7 חלוקת תאDocument24 pagesרזי 7 חלוקת תאBoba BanayNo ratings yet
- 1.6 Cell Division: Essential Idea: Cell Division Is Essential But Must Be ControlledDocument33 pages1.6 Cell Division: Essential Idea: Cell Division Is Essential But Must Be ControlledAnonymous bOMSH4wDgeNo ratings yet
- mitosis-WPS OfficeDocument5 pagesmitosis-WPS OfficeSheila Mae AramanNo ratings yet
- Muhammad Jahanzaib Roll No. 1006 BS Molecular Biology 4 Semester Submitted To:-Sir Muzammil Saeed Developmental BiologyDocument5 pagesMuhammad Jahanzaib Roll No. 1006 BS Molecular Biology 4 Semester Submitted To:-Sir Muzammil Saeed Developmental BiologyBmukulNo ratings yet
- Cell Cycle and Cell DivisionDocument13 pagesCell Cycle and Cell DivisionHeerak SahuNo ratings yet
- Baghdad College of Medical Science (Human Biology)Document9 pagesBaghdad College of Medical Science (Human Biology)علي صلاح مهديNo ratings yet
- HDTD-B-7 - Cell DivisionsDocument43 pagesHDTD-B-7 - Cell DivisionsMariam QaisNo ratings yet
- Cell Cycle and Mitosis Grade 7 BiologyDocument22 pagesCell Cycle and Mitosis Grade 7 BiologyLaszlo ArdaiNo ratings yet
- Bio Topic 3 Student RoomDocument10 pagesBio Topic 3 Student RoomFaizan SanaNo ratings yet
- Stem CellDocument16 pagesStem Celltanishq groverNo ratings yet
- Cell PotencyDocument6 pagesCell PotencyAlexandreNo ratings yet
- Mitosis ThesisDocument5 pagesMitosis Thesisfjnsf5yf100% (2)
- Folio Kerja Kursus Biology SPMDocument29 pagesFolio Kerja Kursus Biology SPMzam_ir100% (2)
- Science 8 Module 4-c3 3Document6 pagesScience 8 Module 4-c3 3Naomi Margaret PetolanNo ratings yet
- Biology ExamDocument17 pagesBiology ExamSai Adrienne RealubitNo ratings yet
- ARDA Embryo 3rd HWDocument3 pagesARDA Embryo 3rd HWSamantha ArdaNo ratings yet
- WEEK7 ModuleLec&LabDocument17 pagesWEEK7 ModuleLec&LabMaricris GatdulaNo ratings yet
- Module 3: Cell Cycle and Cell DivisionDocument8 pagesModule 3: Cell Cycle and Cell DivisionSunidhi ChoudharyNo ratings yet
- Module III - Cell DivisionDocument8 pagesModule III - Cell DivisionSSNo ratings yet
- Module III - Cell DivisionDocument8 pagesModule III - Cell DivisionShivansh SagarNo ratings yet
- Presentation On Cleavage: by Racheal AkpeneDocument28 pagesPresentation On Cleavage: by Racheal AkpeneAkpene RachealNo ratings yet
- 1.3 Cell Division, Diversity and OrganisationDocument12 pages1.3 Cell Division, Diversity and OrganisationMo_Bash1No ratings yet
- Cell DivisionDocument28 pagesCell DivisionPalagiri MadhuNo ratings yet
- Vertebrate Embryonic Cleavage Pattern DeterminationDocument55 pagesVertebrate Embryonic Cleavage Pattern DeterminationtrivikramraoNo ratings yet
- Cell Division Dated 23-08-2013Document24 pagesCell Division Dated 23-08-2013banapte599706No ratings yet
- AP Biology Lab 3 QuestionsDocument3 pagesAP Biology Lab 3 Questionsedwarddame100% (1)
- Mitosis Thesis StatementDocument7 pagesMitosis Thesis Statementzidhelvcf100% (2)
- Chapter 5Document14 pagesChapter 5ingridvirginia2006No ratings yet
- Cell BiologyDocument25 pagesCell BiologyMaricon TubonNo ratings yet
- Cell Division Is The Basis of Reproduction For Every OrganismDocument3 pagesCell Division Is The Basis of Reproduction For Every OrganismAlliah MendozaNo ratings yet
- Pembelahan Embrio (Cleavage) S2 2017Document47 pagesPembelahan Embrio (Cleavage) S2 2017Desy NataliaNo ratings yet
- Essential Idea: Cell Division Is Essential But Must Be ControlledDocument42 pagesEssential Idea: Cell Division Is Essential But Must Be ControlledReeya WhabiNo ratings yet
- Cell CycleDocument111 pagesCell Cyclepreyaaa pascuaNo ratings yet
- Human Chromosomes: An Illustrated Introduction to Human CytogeneticsFrom EverandHuman Chromosomes: An Illustrated Introduction to Human CytogeneticsRating: 5 out of 5 stars5/5 (1)
- Gene Editing, Epigenetic, Cloning and TherapyFrom EverandGene Editing, Epigenetic, Cloning and TherapyRating: 4 out of 5 stars4/5 (1)
- Cell Counting With A Hemocytometer - Easy As 1, 2, 3 - Bitesize Bio PDFDocument10 pagesCell Counting With A Hemocytometer - Easy As 1, 2, 3 - Bitesize Bio PDFShailendra YadavNo ratings yet
- Physiology of Adrenergic ReceptorsDocument3 pagesPhysiology of Adrenergic ReceptorsShailendra YadavNo ratings yet
- Counting Cells in A Hemacytometer - Life TechnologiesDocument1 pageCounting Cells in A Hemacytometer - Life TechnologiesShailendra YadavNo ratings yet
- List of Parasites of Humans - Wikipedia, The Free EncyclopediaDocument12 pagesList of Parasites of Humans - Wikipedia, The Free EncyclopediaShailendra YadavNo ratings yet
- RT PCR PDFDocument2 pagesRT PCR PDFShailendra YadavNo ratings yet
- RT PCR PDFDocument2 pagesRT PCR PDFShailendra YadavNo ratings yet
- λ DNA-HindIII Digest - New England BiolabsDocument3 pagesλ DNA-HindIII Digest - New England BiolabsShailendra YadavNo ratings yet
- Overview of ELISADocument5 pagesOverview of ELISAShailendra YadavNo ratings yet
- How To Write A Scholarship Motivation Letter - ScholarshipPortalDocument3 pagesHow To Write A Scholarship Motivation Letter - ScholarshipPortalShailendra Yadav100% (2)
- Far-Eastern Blotting - Wikipedia, The Free EncyclopediaDocument2 pagesFar-Eastern Blotting - Wikipedia, The Free EncyclopediaShailendra YadavNo ratings yet
- Protein SequencingDocument17 pagesProtein SequencingShiv KumarNo ratings yet
- Gel Electrophoresis - Wikipedia, The Free EncyclopediaDocument10 pagesGel Electrophoresis - Wikipedia, The Free EncyclopediaShailendra YadavNo ratings yet
- Ultracentrifuge - Wikipedia, The Free EncyclopediaDocument3 pagesUltracentrifuge - Wikipedia, The Free EncyclopediaShailendra YadavNo ratings yet
- Klenow Fragment - Wikipedia, The Free EncyclopediaDocument2 pagesKlenow Fragment - Wikipedia, The Free EncyclopediaShailendra YadavNo ratings yet
- Molecular Pathology - Cesium Chloride (CSCL) Density Gradient CentrifugationDocument2 pagesMolecular Pathology - Cesium Chloride (CSCL) Density Gradient CentrifugationShailendra YadavNo ratings yet
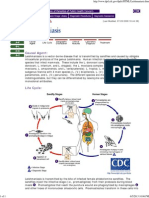
- DPDX - LeishmaniasisDocument1 pageDPDX - LeishmaniasisShailendra YadavNo ratings yet
- Centrosome - Wikipedia, The Free EncyclopediaDocument6 pagesCentrosome - Wikipedia, The Free EncyclopediaShailendra YadavNo ratings yet