You might also like
- California State Bar Order of Discipline of Louis "Skip" Miller For Jury TamperingDocument33 pagesCalifornia State Bar Order of Discipline of Louis "Skip" Miller For Jury TamperingAlan RomeroNo ratings yet
- Motor-Catalog English 2013 PDFDocument80 pagesMotor-Catalog English 2013 PDFILham Dwi PutraNo ratings yet
- NASA: 181330main Jun29colorDocument8 pagesNASA: 181330main Jun29colorNASAdocumentsNo ratings yet
- Transcription, Translation and TransportDocument16 pagesTranscription, Translation and Transportfaria11-6565No ratings yet
- DNA and RNA Viruses Replication-NewDocument48 pagesDNA and RNA Viruses Replication-NewKwirimeNo ratings yet
- Gene ExpressionDocument29 pagesGene Expressionomar khaled العكلNo ratings yet
- Microbial Genetics Lecture - UNTADDocument46 pagesMicrobial Genetics Lecture - UNTADWindy Christine SesaNo ratings yet
- RetroviruseDocument8 pagesRetroviruseMalik RamzanNo ratings yet
- Viral Genetics: Introduction To VirusesDocument22 pagesViral Genetics: Introduction To VirusesMuhammadLukmanulHakimNo ratings yet
- Adeno VirusDocument34 pagesAdeno Virusshikha yadavNo ratings yet
- Corona VirusDocument7 pagesCorona VirusRashmi JhaNo ratings yet
- Alphaviridae PDFDocument1 pageAlphaviridae PDFRahmiati LaoNo ratings yet
- SYBT TranscriptionDocument69 pagesSYBT TranscriptionMeir SabooNo ratings yet
- Microbiology Chapter 4Document45 pagesMicrobiology Chapter 4Milad MovahediNo ratings yet
- Genetics Exam ReviewerDocument9 pagesGenetics Exam ReviewerEysi BarangganNo ratings yet
- Transcription and Regulation of Gene Expression: By: Lyka Marie C. Falcasantos BSN - 1DDocument16 pagesTranscription and Regulation of Gene Expression: By: Lyka Marie C. Falcasantos BSN - 1DRemzAbdullaNo ratings yet
- Structure of RetrovirusDocument12 pagesStructure of RetrovirusMatin Ahmad KhanNo ratings yet
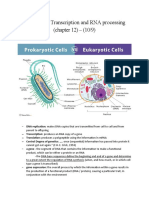
- Transcription and RNA ProcessingDocument38 pagesTranscription and RNA ProcessingRishi Kumar100% (1)
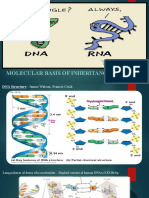
- DNA Structure & InheritanceDocument9 pagesDNA Structure & Inheritancewakeetha cNo ratings yet
- 60855f4d-BIF 14 Gene Expression From Gene To ProteinDocument61 pages60855f4d-BIF 14 Gene Expression From Gene To ProteinMarcus RobinsonNo ratings yet
- EMMRC Lambda Phage 1Document18 pagesEMMRC Lambda Phage 1Polu ChattopadhyayNo ratings yet
- Gene transfer using retro and adeno virusesDocument17 pagesGene transfer using retro and adeno virusesVyshali PingleNo ratings yet
- EXAM II Study Guide (Microbiology)Document25 pagesEXAM II Study Guide (Microbiology)Spencer ThomasNo ratings yet
- Virus Replication StrategyDocument6 pagesVirus Replication StrategyShreyash Raj100% (1)
- Transcriprion ManuscriptDocument14 pagesTranscriprion ManuscriptAsif AhmedNo ratings yet
- On To RetrovirusesDocument4 pagesOn To Retroviruseskep1313No ratings yet
- Lambda Phage Structure and Life CycleDocument21 pagesLambda Phage Structure and Life CycleMagesh RamasamyNo ratings yet
- SummaryDocument47 pagesSummaryDelon van den AkkerNo ratings yet
- Virus Genomes and Its Replication PART 1Document35 pagesVirus Genomes and Its Replication PART 1Mukund VatsNo ratings yet
- PDF 20230417 225731 0000Document27 pagesPDF 20230417 225731 0000Maye JorcaNo ratings yet
- Influenza A lifecycleDocument5 pagesInfluenza A lifecycleTina VargheseNo ratings yet
- Bacterial GeneticsDocument9 pagesBacterial GeneticsKatrine AcostaNo ratings yet
- Virology FinalDocument20 pagesVirology Finalgabrieleciranna23No ratings yet
- DNA to RNA: The Process of TranscriptionDocument10 pagesDNA to RNA: The Process of TranscriptionHardik ManekNo ratings yet
- Mol Basis of InheritanceDocument74 pagesMol Basis of InheritanceNishita BharaliNo ratings yet
- Nurul Aina HakimaDocument23 pagesNurul Aina HakimaAdam AdemNo ratings yet
- Introduction to Virology and Viral ClassificationDocument5 pagesIntroduction to Virology and Viral ClassificationmonchievaleraNo ratings yet
- Microbial GeneticsDocument17 pagesMicrobial Geneticsزين العابدين محمد عويش مشريNo ratings yet
- DR - Fera. Reg Exp Bakteriofaga-virusFeraIbrahim2023Document53 pagesDR - Fera. Reg Exp Bakteriofaga-virusFeraIbrahim2023Zikry SaputraNo ratings yet
- Simian Virus 40 ReplicationDocument10 pagesSimian Virus 40 ReplicationmukundvatsNo ratings yet
- Bacterial Genetics: Ii MbbsDocument28 pagesBacterial Genetics: Ii MbbsmnabrazNo ratings yet
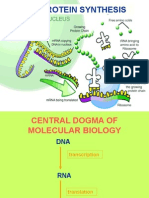
- How Cells Work: The Central DogmaDocument18 pagesHow Cells Work: The Central DogmaJocelyn Diaz AbangNo ratings yet
- Protein SynthesisDocument19 pagesProtein SynthesisminaNo ratings yet
- Laboratorio Biologia MolecolareDocument250 pagesLaboratorio Biologia MolecolareCarIo Alberto Sca0% (1)
- K3 - Central DogmaDocument41 pagesK3 - Central DogmaAditya MuchayatsyahNo ratings yet
- Virologi - DNA Virus ReplicationDocument30 pagesVirologi - DNA Virus ReplicationIvanNo ratings yet
- Tema 7Document11 pagesTema 7Lucía Alba OrdoñezNo ratings yet
- Gene Expression and Population Genetics 2022FDocument114 pagesGene Expression and Population Genetics 2022FGYAN ATTA JOSEPHNo ratings yet
- DNA, RNA, Protein-Gene ExpressionDocument18 pagesDNA, RNA, Protein-Gene ExpressionFahim AbidNo ratings yet
- Microbiology With Diseases by Body System 4th Edition Bauman Solutions ManualDocument9 pagesMicrobiology With Diseases by Body System 4th Edition Bauman Solutions Manualzacharymeliora0h86100% (31)
- CRISPR CAS 9 Is Found inDocument12 pagesCRISPR CAS 9 Is Found inHamza KhanNo ratings yet
- Microbiology With Diseases by Body System 4th Edition Bauman Solutions ManualDocument39 pagesMicrobiology With Diseases by Body System 4th Edition Bauman Solutions Manualcharles90br100% (13)
- Microbiology With Diseases by Body System 4Th Edition Bauman Solutions Manual Full Chapter PDFDocument30 pagesMicrobiology With Diseases by Body System 4Th Edition Bauman Solutions Manual Full Chapter PDFlloydkieran71epfi100% (10)
- Bio Project 2019Document7 pagesBio Project 2019Ericom On RiqcousNo ratings yet
- Retrovirus Replication by Bhuvanesh KalalDocument29 pagesRetrovirus Replication by Bhuvanesh Kalalbhuvanesh Sukhlal100% (2)
- Gene ExpressionDocument19 pagesGene Expressionkvicto100% (1)
- Plant Transformation MethodsDocument43 pagesPlant Transformation MethodsaaasidNo ratings yet
- Study-Questions Module-3 MCB104Document3 pagesStudy-Questions Module-3 MCB104Ano NymousNo ratings yet
- Anatomy of A GeneDocument33 pagesAnatomy of A GenemskikiNo ratings yet
- Presentasi MikrobioDocument28 pagesPresentasi MikrobioKuswati FaisolNo ratings yet
- Lecture 2 - Transcription and RNA ProcessingDocument22 pagesLecture 2 - Transcription and RNA ProcessingErika KulićNo ratings yet
- Structural Biology of HIVDocument31 pagesStructural Biology of HIVLaura TapiaNo ratings yet
- 1967 Painting Israeli VallejoDocument1 page1967 Painting Israeli VallejoMiloš CiniburkNo ratings yet
- Duttaphrynus Melanostictus,: Errata VersionDocument11 pagesDuttaphrynus Melanostictus,: Errata Versionutama 3002No ratings yet
- 7) Set 3 Bi PT3 (Answer) PDFDocument4 pages7) Set 3 Bi PT3 (Answer) PDFTing ShiangNo ratings yet
- Anne Frank Halloween Costume 2Document5 pagesAnne Frank Halloween Costume 2ive14_No ratings yet
- Guide To Kulchur 1938Document374 pagesGuide To Kulchur 1938Jonathan CarsonNo ratings yet
- You Write, It Types!: Quick Start GuideDocument21 pagesYou Write, It Types!: Quick Start Guidejean michelNo ratings yet
- 6 Fsiqiatria-1524041346Document48 pages6 Fsiqiatria-1524041346მირანდა გიორგაშვილიNo ratings yet
- Written Work 1 Q1 Science 10Document6 pagesWritten Work 1 Q1 Science 10JOEL MONTERDENo ratings yet
- SDM - Session 6Document21 pagesSDM - Session 6Rohith NairNo ratings yet
- Packex IndiaDocument12 pagesPackex IndiaSam DanNo ratings yet
- UITM Faculty of Business Entrepreneurship Rubric Social Media PortfolioDocument9 pagesUITM Faculty of Business Entrepreneurship Rubric Social Media PortfolioShamsyul AriffinNo ratings yet
- Spreadsheet and Presentation Skills SyllabusDocument4 pagesSpreadsheet and Presentation Skills SyllabusGbox CTCNo ratings yet
- SCM Software Selection and EvaluationDocument3 pagesSCM Software Selection and EvaluationBhuwneshwar PandayNo ratings yet
- The Oz DietDocument5 pagesThe Oz Dietkaren_wilkesNo ratings yet
- MarketNexus Editor: Teri Buhl Character LetterDocument2 pagesMarketNexus Editor: Teri Buhl Character LetterTeri BuhlNo ratings yet
- Perlman Janice, The Myth of Marginality RevisitDocument36 pagesPerlman Janice, The Myth of Marginality RevisitLuisa F. RodriguezNo ratings yet
- Mora Solvendi (Delay of The Debtor)Document11 pagesMora Solvendi (Delay of The Debtor)John Paul100% (1)
- Criteria For Judging MR and Ms UNDocument9 pagesCriteria For Judging MR and Ms UNRexon ChanNo ratings yet
- Types of Companies Classified by Incorporation, Membership, Liability and ControlDocument11 pagesTypes of Companies Classified by Incorporation, Membership, Liability and ControlPrasad BulbuleNo ratings yet
- Lesson 5 Classifications of CommunicationDocument48 pagesLesson 5 Classifications of CommunicationRovenick SinggaNo ratings yet
- 2013 SmartBUS Home Automation Product Catalogue English v.1.0Document108 pages2013 SmartBUS Home Automation Product Catalogue English v.1.0Smart-G4100% (3)
- Target products to meet 20% demandDocument12 pagesTarget products to meet 20% demandAlma Dela PeñaNo ratings yet
- 2689 26489 1 PBDocument9 pages2689 26489 1 PBAntonio EspañaNo ratings yet
- Policy Based Routing On Fortigate FirewallDocument2 pagesPolicy Based Routing On Fortigate FirewalldanNo ratings yet
- Joyce2016 PDFDocument13 pagesJoyce2016 PDFAffan ArrizqiNo ratings yet
- Teresita Dio Versus STDocument2 pagesTeresita Dio Versus STmwaike100% (1)
- Protective & Marine Coatings: Hi-Solids Alkyd Metal PrimerDocument4 pagesProtective & Marine Coatings: Hi-Solids Alkyd Metal PrimerAna CabreraNo ratings yet