You might also like
- The Mars ForceDocument249 pagesThe Mars Forceridikitty100% (2)
- Identification and Evaluation of Probiotic Potential in Yeast Strains from KefirDocument7 pagesIdentification and Evaluation of Probiotic Potential in Yeast Strains from KefirDaniel InzunzaNo ratings yet
- Second Quarterly Examination Math 9Document2 pagesSecond Quarterly Examination Math 9Mark Kiven Martinez94% (16)
- Chronological OrderDocument5 pagesChronological OrderDharWin d'Wing-Wing d'AriestBoyzNo ratings yet
- Daily Assessment RecordDocument4 pagesDaily Assessment Recordapi-342236522100% (2)
- Oracle Unified Method (OUM) White Paper - Oracle's Full Lifecycle Method For Deploying Oracle-Based Business Solutions - GeneralDocument17 pagesOracle Unified Method (OUM) White Paper - Oracle's Full Lifecycle Method For Deploying Oracle-Based Business Solutions - GeneralAndreea Mirosnicencu100% (1)
- Business Analysis FoundationsDocument39 pagesBusiness Analysis FoundationsPriyankaNo ratings yet
- The Stressing Life of Lactobacillus Delbrueckii Subsp. Bulgaricus in Soy Milk (2022)Document15 pagesThe Stressing Life of Lactobacillus Delbrueckii Subsp. Bulgaricus in Soy Milk (2022)Vũ Quyết ThắngNo ratings yet
- Production of A Novel Probiotic YogurtDocument10 pagesProduction of A Novel Probiotic YogurtLilíNo ratings yet
- 1 s2.0 S0958694619300317 MainDocument10 pages1 s2.0 S0958694619300317 Mainrevathimr2311No ratings yet
- 1-S2.0-S0740002007001001-Main (Incorporation of Citrus Fibers in Fermented MilkDocument9 pages1-S2.0-S0740002007001001-Main (Incorporation of Citrus Fibers in Fermented MilkEddy WibowoNo ratings yet
- LWT - Food Science and Technology 116 (2019) 108476Document8 pagesLWT - Food Science and Technology 116 (2019) 108476Đức TrầnNo ratings yet
- 1 s2.0 S1756464616300986 MainDocument11 pages1 s2.0 S1756464616300986 MainSarfrazNo ratings yet
- Food Research International: C.P. Champagne, J. Green-Johnson, Y. Raymond, J. Barrette, N. BuckleyDocument10 pagesFood Research International: C.P. Champagne, J. Green-Johnson, Y. Raymond, J. Barrette, N. BuckleyNaufal BaghirNo ratings yet
- Valli 2015Document7 pagesValli 2015Katalin BodorNo ratings yet
- 1 s2.0 S0023643818303591 MainDocument8 pages1 s2.0 S0023643818303591 MainSarfrazNo ratings yet
- Yaourt Foodand Health 2016Document10 pagesYaourt Foodand Health 2016Patrick ParraNo ratings yet
- Impact of Banana Puree On Viability of P PDFDocument8 pagesImpact of Banana Puree On Viability of P PDFAndreea FlorentinaNo ratings yet
- New Perspectives in Fermented Dairy Products and Their Health RelevanceDocument11 pagesNew Perspectives in Fermented Dairy Products and Their Health RelevanceAndrea Osorio AlturoNo ratings yet
- MLJ 72 201 enDocument12 pagesMLJ 72 201 enmilu1312No ratings yet
- Topic 18Document15 pagesTopic 18Nguyen Minh TrongNo ratings yet
- J Aoas 2020 03 001Document10 pagesJ Aoas 2020 03 001Raksa SaremNo ratings yet
- Growth of Probiotic in Soy Yogurt Formulation PDFDocument8 pagesGrowth of Probiotic in Soy Yogurt Formulation PDFLenaldy Nuari GarnokoNo ratings yet
- Lazo-Vélez, Chuck-Hernandez, Serna-Saldívar - 2015 - Evaluation of The Functionality of Five Different Soybean Proteins in Yeast-LeaveneDocument7 pagesLazo-Vélez, Chuck-Hernandez, Serna-Saldívar - 2015 - Evaluation of The Functionality of Five Different Soybean Proteins in Yeast-LeaveneRocy FernándezNo ratings yet
- Evaluation of The Effect of Supplementing Fermented Milk With Quinoa Flour On Probiotic ActivityDocument9 pagesEvaluation of The Effect of Supplementing Fermented Milk With Quinoa Flour On Probiotic Activitywilliam LlantuyNo ratings yet
- Novel Insights Into The Microbiology of Fermented Dairy FoodsDocument7 pagesNovel Insights Into The Microbiology of Fermented Dairy FoodsRuben SerraNo ratings yet
- Health Benefits of Yogurt Consumption A ReviewDocument13 pagesHealth Benefits of Yogurt Consumption A ReviewDark PrincessNo ratings yet
- 607 59912 1 PBDocument12 pages607 59912 1 PBRobi Amizar SikumbangNo ratings yet
- Novel insights into microbiology of fermented dairyDocument9 pagesNovel insights into microbiology of fermented dairyoscarandresNo ratings yet
- Idoneidad de La Fruta de Pitaya Fermentada Por Cepas de LAB de Masa MadreDocument9 pagesIdoneidad de La Fruta de Pitaya Fermentada Por Cepas de LAB de Masa MadreELIANA AÍDA LÓPEZ CORONADONo ratings yet
- 2023 Metagenomic Analysis Antibacterial Activity Kefir MicroorganismsDocument17 pages2023 Metagenomic Analysis Antibacterial Activity Kefir MicroorganismsHumberto MadeiraNo ratings yet
- 1 s2.0 S2666833521000381 MainDocument11 pages1 s2.0 S2666833521000381 Mainiñaki zamora sevillaNo ratings yet
- Bacillus CoagulansDocument14 pagesBacillus CoagulansMª Verónica B.No ratings yet
- Probiotic Fermented Milks Present and FutureDocument12 pagesProbiotic Fermented Milks Present and FutureJuan Jose Escobar P.No ratings yet
- Tamer Prob - Dond.Document9 pagesTamer Prob - Dond.Nicole PazmiñoNo ratings yet
- Developing probiotic buttermilk on a semi-industrial scaleDocument8 pagesDeveloping probiotic buttermilk on a semi-industrial scaleUyên SandyNo ratings yet
- Her Verte Tal 2017Document12 pagesHer Verte Tal 2017Abhishek BanerjeeNo ratings yet
- Hygienic and Microbial Quality of YoghurtDocument4 pagesHygienic and Microbial Quality of YoghurtRobert RossNo ratings yet
- Viability and Activity of Bifidobacteria in Yogurt Containing Fructooligosaccharide During Refrigerated StorageDocument10 pagesViability and Activity of Bifidobacteria in Yogurt Containing Fructooligosaccharide During Refrigerated StorageLoredana Veronica ZalischiNo ratings yet
- Evaluation of The Rheological, Textural, Microstructural andDocument11 pagesEvaluation of The Rheological, Textural, Microstructural andJorge RamirezNo ratings yet
- Heenan 2002Document6 pagesHeenan 2002Jorge SaenzNo ratings yet
- Research Proposal: Lactic Acid Bacteria and Development of Microbial AgentsDocument3 pagesResearch Proposal: Lactic Acid Bacteria and Development of Microbial AgentsNaveed AhmadNo ratings yet
- Jayalalitha, V., Palani Dorai. R., Dhanalakshmi, B., Elango, A. and Naresh Kumar CDocument4 pagesJayalalitha, V., Palani Dorai. R., Dhanalakshmi, B., Elango, A. and Naresh Kumar CThasya TamaraNo ratings yet
- Whole ThesisDocument201 pagesWhole ThesisNache GeorgianNo ratings yet
- Aj 1Document11 pagesAj 1Trần Tấn TàiNo ratings yet
- Naturally Fermented Milk and Its Therapeutic Potential in The Treatment of Inflammatory Intestinal DisordersDocument17 pagesNaturally Fermented Milk and Its Therapeutic Potential in The Treatment of Inflammatory Intestinal DisordersRuby SakaNo ratings yet
- Development and Physico Chemical Analysis of Probiotic Vegan Yogurt Using Artocarpus Heterophyllus Jackfruit Seed and Cocos Nucifera Coconut MilkDocument5 pagesDevelopment and Physico Chemical Analysis of Probiotic Vegan Yogurt Using Artocarpus Heterophyllus Jackfruit Seed and Cocos Nucifera Coconut MilkResearch ParkNo ratings yet
- 0101 2061 Cta fst24818 (Importa)Document10 pages0101 2061 Cta fst24818 (Importa)GRACIELANo ratings yet
- (Articulo 6) PDFDocument10 pages(Articulo 6) PDFmagaNo ratings yet
- Microbiology of Fermented Dairy Products PDFDocument12 pagesMicrobiology of Fermented Dairy Products PDFJura SenykNo ratings yet
- 904 Guetouache Et AlDocument9 pages904 Guetouache Et AlsamirNo ratings yet
- Boosting milk production with biodiesel byproductsDocument11 pagesBoosting milk production with biodiesel byproductsLarissa InacioNo ratings yet
- Các Lo I Đư NG Dùng Cho LABDocument15 pagesCác Lo I Đư NG Dùng Cho LABThương PhanNo ratings yet
- Microbial Biotechnology - 2022 - Harper - Fermentation of Plant Based Dairy Alternatives by Lactic Acid BacteriaDocument18 pagesMicrobial Biotechnology - 2022 - Harper - Fermentation of Plant Based Dairy Alternatives by Lactic Acid BacteriaMartin Oscar SanchezNo ratings yet
- Microbial Quality and Safety of Milk and Milk Products in TheDocument37 pagesMicrobial Quality and Safety of Milk and Milk Products in TheWilson SilvaNo ratings yet
- S. Langa, A. Peirot En, P. Gaya, C. Escudero, E. Rodríguez-Mínguez, J.M. Landete, J.L. Arqu EsDocument8 pagesS. Langa, A. Peirot En, P. Gaya, C. Escudero, E. Rodríguez-Mínguez, J.M. Landete, J.L. Arqu EsĐức TrầnNo ratings yet
- Semisweet Chocolate As A Vehicle...Document8 pagesSemisweet Chocolate As A Vehicle...Kartika KeksiNo ratings yet
- International Dairy JournalDocument9 pagesInternational Dairy JournalYair Fernandez SanchezNo ratings yet
- Development and Evaluation of Functional Biscuits From Underutilised Crops of LadakhDocument11 pagesDevelopment and Evaluation of Functional Biscuits From Underutilised Crops of Ladakhanwar hussainNo ratings yet
- Mycotoxins in Milk Occurrence and Evaluation of CeDocument16 pagesMycotoxins in Milk Occurrence and Evaluation of CeBorja Muñoz SolanoNo ratings yet
- 1 s2.0 S0023643819301732 MainDocument7 pages1 s2.0 S0023643819301732 MainSarfrazNo ratings yet
- LWT - Food Science and TechnologyDocument8 pagesLWT - Food Science and TechnologyRominaNo ratings yet
- Manuscript Almond Version AutorDocument40 pagesManuscript Almond Version AutorLoredana Veronica ZalischiNo ratings yet
- Development of sprouted wheat based probiotic beverageDocument8 pagesDevelopment of sprouted wheat based probiotic beverageSipend AnatomiNo ratings yet
- Evaluation of Physicochemical Traits, Sensory Properties and Survival of Lactobacillus Casei in Fermented Soy-Based Ice CreamDocument7 pagesEvaluation of Physicochemical Traits, Sensory Properties and Survival of Lactobacillus Casei in Fermented Soy-Based Ice CreampriyankaNo ratings yet
- Impact of Various Types of Heat Processing On The Energy and Nutritional Values of Goose Breast MeatDocument14 pagesImpact of Various Types of Heat Processing On The Energy and Nutritional Values of Goose Breast MeatNaveed AkramNo ratings yet
- MNS Agriculture Post Harvest ManagementDocument13 pagesMNS Agriculture Post Harvest ManagementNaveed AkramNo ratings yet
- LWT - Food Science and Technology 157 (2022) 113103Document9 pagesLWT - Food Science and Technology 157 (2022) 113103Naveed AkramNo ratings yet
- LWT - Food Science and Technology 157 (2022) 113103Document9 pagesLWT - Food Science and Technology 157 (2022) 113103Naveed AkramNo ratings yet
- 1 s2.0 S002203022101122X MainDocument11 pages1 s2.0 S002203022101122X MainNaveed AkramNo ratings yet
- Food Hydrocolloids For Health: Salvia HispanicaDocument5 pagesFood Hydrocolloids For Health: Salvia HispanicaNaveed AkramNo ratings yet
- Service Parts List: 54-26-0005 2551-20 M12™ FUEL™ SURGE™ 1/4" Hex Hydraulic Driver K42ADocument2 pagesService Parts List: 54-26-0005 2551-20 M12™ FUEL™ SURGE™ 1/4" Hex Hydraulic Driver K42AAmjad AlQasrawi100% (1)
- 19 Unpriced BOM, Project & Manpower Plan, CompliancesDocument324 pages19 Unpriced BOM, Project & Manpower Plan, CompliancesAbhay MishraNo ratings yet
- ProjectDocument86 pagesProjectrajuNo ratings yet
- An Overview of The FUPLA 2 Tools: Project DatabaseDocument2 pagesAn Overview of The FUPLA 2 Tools: Project DatabaseJulio Cesar Rojas SaavedraNo ratings yet
- How Do I Prepare For Public Administration For IAS by Myself Without Any Coaching? Which Books Should I Follow?Document3 pagesHow Do I Prepare For Public Administration For IAS by Myself Without Any Coaching? Which Books Should I Follow?saiviswanath0990100% (1)
- Barcelona Smart City TourDocument44 pagesBarcelona Smart City TourPepe JeansNo ratings yet
- CURRICULUM AUDIT: GRADE 7 MATHEMATICSDocument5 pagesCURRICULUM AUDIT: GRADE 7 MATHEMATICSjohnalcuinNo ratings yet
- The Interview: P F T IDocument14 pagesThe Interview: P F T IkkkkccccNo ratings yet
- Dyna 2000 LiteDocument2 pagesDyna 2000 LiteRNKNo ratings yet
- GD&T WIZ Tutor Covers The Vast Breadth of Geometric Dimensioning and Tolerancing Without Compromising On The Depth. The Topics Covered AreDocument1 pageGD&T WIZ Tutor Covers The Vast Breadth of Geometric Dimensioning and Tolerancing Without Compromising On The Depth. The Topics Covered AreVinay ManjuNo ratings yet
- Geometric Sculpture From 72 PencilsDocument5 pagesGeometric Sculpture From 72 PencilsHugo Iván Gonzalez CruzNo ratings yet
- Corporate Governance in SMEsDocument18 pagesCorporate Governance in SMEsSana DjaanineNo ratings yet
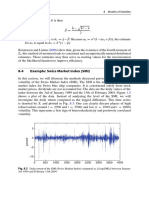
- 8.4 Example: Swiss Market Index (SMI) : 188 8 Models of VolatilityDocument3 pages8.4 Example: Swiss Market Index (SMI) : 188 8 Models of VolatilityNickesh ShahNo ratings yet
- Abstract Photo CompositionDocument2 pagesAbstract Photo Compositionapi-260853196No ratings yet
- Chapter 2 RM - Research ProcessDocument46 pagesChapter 2 RM - Research ProcesseyasuNo ratings yet
- Organic Chem Diels-Alder Reaction LabDocument9 pagesOrganic Chem Diels-Alder Reaction LabPryanka BalleyNo ratings yet
- Galambos 1986Document18 pagesGalambos 1986gcoNo ratings yet
- List of household items for relocationDocument4 pagesList of household items for relocationMADDYNo ratings yet
- ElectricalDocument30 pagesElectricalketerNo ratings yet
- PedigreesDocument5 pagesPedigreestpn72hjg88No ratings yet
- TILE FIXING GUIDEDocument1 pageTILE FIXING GUIDEStavros ApostolidisNo ratings yet
- Operating Manual: Please Read These Operating Instructions Before Using Your FreedomchairDocument24 pagesOperating Manual: Please Read These Operating Instructions Before Using Your FreedomchairNETHYA SHARMANo ratings yet
- Common Pesticides in AgricultureDocument6 pagesCommon Pesticides in AgricultureBMohdIshaqNo ratings yet
- CHEMICAL ANALYSIS OF WATER SAMPLEDocument5 pagesCHEMICAL ANALYSIS OF WATER SAMPLEAiron Fuentes EresNo ratings yet