You might also like
- A Global Phylogeny of Apple Snails: Gondwanan Origin, Generic Relationships, and The Influence of Outgroup Choice (Caenogastropoda: Ampullariidae)Document16 pagesA Global Phylogeny of Apple Snails: Gondwanan Origin, Generic Relationships, and The Influence of Outgroup Choice (Caenogastropoda: Ampullariidae)SandraJustinoNo ratings yet
- Momotus Revision GeneroDocument47 pagesMomotus Revision GeneroYoelis Yepes PerezNo ratings yet
- Tmp2a27 TMPDocument15 pagesTmp2a27 TMPFrontiersNo ratings yet
- Key and Phylogeny Paratetrapedia, Aguiar & Melo 2010Document92 pagesKey and Phylogeny Paratetrapedia, Aguiar & Melo 2010Nicolas David Gutierrez RubianoNo ratings yet
- Deforeletal 2021melanophryniscusDocument27 pagesDeforeletal 2021melanophryniscusjose.1711717678No ratings yet
- Ramos Etal 2022 Evolution Andrenine BeesDocument11 pagesRamos Etal 2022 Evolution Andrenine BeesTayaneFebroneNo ratings yet
- Interacciones de BatoideosDocument22 pagesInteracciones de BatoideosALHONDRA HORTENCIA GOMEZ SANCHEZNo ratings yet
- Phylogeny of The NymphalidaeDocument21 pagesPhylogeny of The Nymphalidaelv acNo ratings yet
- Doria Et Al 2008 MenispermaceaeDocument20 pagesDoria Et Al 2008 Menispermaceaefabianyht1978No ratings yet
- Ammerman, L., Lee, D. y Tipp, M. (2012) .Document17 pagesAmmerman, L., Lee, D. y Tipp, M. (2012) .lina maaria lopezNo ratings yet
- The Never Ending Story: Multigene Approaches To The Phylogeny of AmarillidaceaeDocument14 pagesThe Never Ending Story: Multigene Approaches To The Phylogeny of Amarillidaceaeדר בן-נתןNo ratings yet
- Herrera Et Al - 2011 - Paleocene Menispermaceae Endocarps From Colombia & WyomingDocument15 pagesHerrera Et Al - 2011 - Paleocene Menispermaceae Endocarps From Colombia & Wyomingfabianyht1978No ratings yet
- Cecca F. & Alii 2006 - Evolution of The Tithonian Ammonite SemiformicerasDocument12 pagesCecca F. & Alii 2006 - Evolution of The Tithonian Ammonite Semiformiceraschin dasNo ratings yet
- 0102-33062016abb0401 InddDocument9 pages0102-33062016abb0401 InddAlén MirandaNo ratings yet
- Filogenia LampropeltisDocument11 pagesFilogenia LampropeltisDavid BrizNo ratings yet
- A Review of The Nearctic Jumping SpidersDocument12 pagesA Review of The Nearctic Jumping SpidersGenaroNo ratings yet
- Turco Bologna Meloidae Gen N 2008Document14 pagesTurco Bologna Meloidae Gen N 2008Jean-Bernard HuchetNo ratings yet
- Molecular Phylogeny of Mugilidae Fishes RevisedDocument15 pagesMolecular Phylogeny of Mugilidae Fishes RevisedVineet DubeyNo ratings yet
- 9-Fernandez Et Al. 2017 Azorella-Laretia-Mulinum Phylogeny-MPEDocument21 pages9-Fernandez Et Al. 2017 Azorella-Laretia-Mulinum Phylogeny-MPEMartina FernandezNo ratings yet
- Evolution - February 1996 - Shaffer - THE POLYTYPIC SPECIES REVISITED GENETIC DIFFERENTIATION AND MOLECULAR PHYLOGENETICSDocument17 pagesEvolution - February 1996 - Shaffer - THE POLYTYPIC SPECIES REVISITED GENETIC DIFFERENTIATION AND MOLECULAR PHYLOGENETICSmendozar787No ratings yet
- American J of Botany - 2001 - Clausing - Molecular Phylogenetics of Melastomataceae and Memecylaceae Implications ForDocument13 pagesAmerican J of Botany - 2001 - Clausing - Molecular Phylogenetics of Melastomataceae and Memecylaceae Implications Formateus silvaNo ratings yet
- Joseph 2004Document14 pagesJoseph 2004Eduardo Zavala B.No ratings yet
- Van Der Hoof 2003 Phylogenia HystiotusDocument63 pagesVan Der Hoof 2003 Phylogenia Hystiotusruthmcontreras88No ratings yet
- Lima Et Al. 2014 Cratosmilax JacksoniDocument16 pagesLima Et Al. 2014 Cratosmilax JacksoniFlaviana LimaNo ratings yet
- Boj 12053Document18 pagesBoj 12053SWAPNIL CHAURASIYANo ratings yet
- Molecular Phylogenetics and Evolution: Sciverse SciencedirectDocument11 pagesMolecular Phylogenetics and Evolution: Sciverse SciencedirectVermilion~No ratings yet
- Pinos Pinioneros2000Document14 pagesPinos Pinioneros2000Coraline JonesNo ratings yet
- Gomez-Acevedo 2010 Neotropical Mutualism Between Acacia and Pseudomyrmex Phylogeny and Divergence TimesDocument16 pagesGomez-Acevedo 2010 Neotropical Mutualism Between Acacia and Pseudomyrmex Phylogeny and Divergence TimesTheChaoticFlameNo ratings yet
- Exploring Phenotypic Plasticity and Biogeography in Emerald MothsDocument11 pagesExploring Phenotypic Plasticity and Biogeography in Emerald Mothsluras.zapatalNo ratings yet
- 230953-Article Text-560517-1-10-20220831Document14 pages230953-Article Text-560517-1-10-20220831fireworkNo ratings yet
- Bravo Et Al. 2012Document7 pagesBravo Et Al. 2012qplygxlj9No ratings yet
- Molecular Phylogenetics and EvolutionDocument11 pagesMolecular Phylogenetics and EvolutionOanaNo ratings yet
- Simon Et Al 2011 American Journal of BotanyDocument21 pagesSimon Et Al 2011 American Journal of BotanyTaline PinheiroNo ratings yet
- Campbell E - Otomanguean Historical Ling - 240325 - 213315Document40 pagesCampbell E - Otomanguean Historical Ling - 240325 - 213315jcgallardhoNo ratings yet
- Comparative+mitogenome+phylogeography+of+two+anteater+genera+ (Tamandua+and+Myrmecophaga +Myrmecophagidae,+Xenarthra) +evidence+of+discrepant+evolutionary+traitsDocument23 pagesComparative+mitogenome+phylogeography+of+two+anteater+genera+ (Tamandua+and+Myrmecophaga +Myrmecophagidae,+Xenarthra) +evidence+of+discrepant+evolutionary+traitsManuel RuizNo ratings yet
- Phylogenetic - Analysis - of - The - Lythraceae - 20160720 29388 1qgm2lw With Cover Page v2Document24 pagesPhylogenetic - Analysis - of - The - Lythraceae - 20160720 29388 1qgm2lw With Cover Page v2Lina María Ospina RojasNo ratings yet
- Abello Et Al 2015 Zootaxa 3937Document14 pagesAbello Et Al 2015 Zootaxa 3937mioclaenusNo ratings yet
- Lobster Phyl Ogen yDocument11 pagesLobster Phyl Ogen ykirandasari143No ratings yet
- Primate Phylogeny Morphology Vs Molecular ResultsDocument53 pagesPrimate Phylogeny Morphology Vs Molecular ResultsCosette JavðóttirNo ratings yet
- Mohammadin 2017 Anatolian Origins and DiversificatiDocument13 pagesMohammadin 2017 Anatolian Origins and DiversificatisalarNo ratings yet
- Fossil Evidence and The Origin of Bats: Gregg F. Gunnell and Nancy B. SimmonsDocument38 pagesFossil Evidence and The Origin of Bats: Gregg F. Gunnell and Nancy B. SimmonsRenz Junyll ApigoNo ratings yet
- Alonso - Crespo. Taxonomic and Nomenclatural Notes On South American Taxa ...Document14 pagesAlonso - Crespo. Taxonomic and Nomenclatural Notes On South American Taxa ...Biblioteca ImbivNo ratings yet
- Tithaeidae New Species From Mindanao Schmidt Et Al 2019Document15 pagesTithaeidae New Species From Mindanao Schmidt Et Al 2019JOHN JERALD VILLAMANCANo ratings yet
- Taxonomic Revision of The Genus CrassicrusDocument32 pagesTaxonomic Revision of The Genus CrassicrusJosé Arturo S. TéllezNo ratings yet
- Bertelli Et Al. 2014 - Phylogenetic Interrelationships of Living and Extinct Tinamidae From The New WorldDocument40 pagesBertelli Et Al. 2014 - Phylogenetic Interrelationships of Living and Extinct Tinamidae From The New WorldJohselmCantoNo ratings yet
- Flora of SurinameDocument147 pagesFlora of SurinameMolineNo ratings yet
- Jenny 2000 Permophiles 37Document10 pagesJenny 2000 Permophiles 37Gerard StampfliNo ratings yet
- EditorialstinglessbeesDocument6 pagesEditorialstinglessbeesManap TriantoNo ratings yet
- Molecular Phylogeny of DipetrocarpaceaeDocument8 pagesMolecular Phylogeny of DipetrocarpaceaedugongNo ratings yet
- Bromeliad EvolutionDocument25 pagesBromeliad EvolutionLorena VillalobosNo ratings yet
- JNatHist 2018 Cox EtalDocument44 pagesJNatHist 2018 Cox EtalherpmxNo ratings yet
- Wallach Epictia PaperDocument160 pagesWallach Epictia PaperherpmxNo ratings yet
- Marcelo Dias Machado Vianna-Filho Andrea Ferreira Da CostaDocument13 pagesMarcelo Dias Machado Vianna-Filho Andrea Ferreira Da CostaMarcelo ViannaNo ratings yet
- A New Species of Monodelphis (Didelphimorphia: Didelphidae) From Southeastern PeruDocument8 pagesA New Species of Monodelphis (Didelphimorphia: Didelphidae) From Southeastern PeruJulio César Chávez GalarzaNo ratings yet
- Wang & Carranza (2008) - Conepatus From E-Pliocene of Guanajuato (BLANCAN) PDFDocument22 pagesWang & Carranza (2008) - Conepatus From E-Pliocene of Guanajuato (BLANCAN) PDFArturo Palma RamírezNo ratings yet
- Abzhanov Kaufman 2000 EvolutionDocument14 pagesAbzhanov Kaufman 2000 EvolutionDavid CarrozzoNo ratings yet
- Areta Et Al 2023 - Amaurospiza PDFDocument18 pagesAreta Et Al 2023 - Amaurospiza PDFMajo TorallaNo ratings yet
- Phylogenetic Relationships of Calomys Sorellus Complex (Rodentia: Cricetidae), With The Description of Two New SpeciesDocument24 pagesPhylogenetic Relationships of Calomys Sorellus Complex (Rodentia: Cricetidae), With The Description of Two New SpeciesRudy ConchaNo ratings yet
- Insectos Del Cenozoico en La ArgentinaDocument9 pagesInsectos Del Cenozoico en La ArgentinaDavid ArenasNo ratings yet
- Australian Lauxaniid Flies: Revision of the Australian Species of Homoneura van der Wulp, Trypetisoma Malloch, and Allied Genera (Diptera : Lauxaniidae)From EverandAustralian Lauxaniid Flies: Revision of the Australian Species of Homoneura van der Wulp, Trypetisoma Malloch, and Allied Genera (Diptera : Lauxaniidae)No ratings yet
- 2021.sayustietal - StinglessbeesSulawesimorphologyneststructureandmolecular JARDocument16 pages2021.sayustietal - StinglessbeesSulawesimorphologyneststructureandmolecular JARManap TriantoNo ratings yet
- 2023 Vanessa Almasdi EfektifitasPenggunaan7no2Document10 pages2023 Vanessa Almasdi EfektifitasPenggunaan7no2Manap TriantoNo ratings yet
- 1 PBDocument56 pages1 PBManap TriantoNo ratings yet
- 1048-Article Text-4531-1-10-20230310Document18 pages1048-Article Text-4531-1-10-20230310Manap TriantoNo ratings yet
- Dear Editor-in-Chief,: Covering LetterDocument2 pagesDear Editor-in-Chief,: Covering LetterManap TriantoNo ratings yet
- Wordwall As A Learning Media To Increase Students' Reading InterestDocument9 pagesWordwall As A Learning Media To Increase Students' Reading Interestrefiam2016No ratings yet
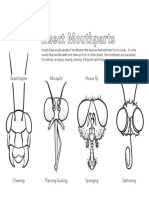
- Colouring Sheet Insect MouthpartsDocument1 pageColouring Sheet Insect MouthpartsManap TriantoNo ratings yet
- Study On The Beekeeping Situation The Level of BeeDocument6 pagesStudy On The Beekeeping Situation The Level of BeeManap TriantoNo ratings yet
- Invoice - Ix 1 1888Document1 pageInvoice - Ix 1 1888Manap TriantoNo ratings yet
- Hines2005 - FIX 4Document14 pagesHines2005 - FIX 4Manap TriantoNo ratings yet
- Letter of Acceptance - IX-1-1888Document1 pageLetter of Acceptance - IX-1-1888Manap TriantoNo ratings yet
- EditorialstinglessbeesDocument6 pagesEditorialstinglessbeesManap TriantoNo ratings yet
- Preen1997 Rawan - FIX 6Document25 pagesPreen1997 Rawan - FIX 6Manap TriantoNo ratings yet
- Lanyon2003 - FIX 5Document13 pagesLanyon2003 - FIX 5Manap TriantoNo ratings yet
- Attachment - FIX 3Document13 pagesAttachment - FIX 3Manap TriantoNo ratings yet
- Camargo and Roubik 2005Document15 pagesCamargo and Roubik 2005Manap TriantoNo ratings yet
- Building Phylogenetic Trees From Molecular Data With MEGA: Molecular Biology and Evolution March 2013Document8 pagesBuilding Phylogenetic Trees From Molecular Data With MEGA: Molecular Biology and Evolution March 2013Manap TriantoNo ratings yet
- Awra Amba RJ 300612 EN BDDocument86 pagesAwra Amba RJ 300612 EN BDTop ListorsNo ratings yet
- Virtue Ethics: Deficiency ExcessDocument1 pageVirtue Ethics: Deficiency ExcesschristianNo ratings yet
- Institute of Southern Punjab Multan: Syed Zohair Quain Haider Lecturer ISP MultanDocument47 pagesInstitute of Southern Punjab Multan: Syed Zohair Quain Haider Lecturer ISP MultanCh MaaanNo ratings yet
- TRANS Finals NSTPDocument6 pagesTRANS Finals NSTPStephanieNo ratings yet
- Directorate General of Drug Administration: SL Name of The Pharmaceutical Address Location Licence No. Present StatusDocument2 pagesDirectorate General of Drug Administration: SL Name of The Pharmaceutical Address Location Licence No. Present StatusAnamika SahaNo ratings yet
- Mes 038Document9 pagesMes 038MimiNo ratings yet
- My Side of The Mountain PDFDocument61 pagesMy Side of The Mountain PDF12333% (3)
- Easy Way To Spot Elliott Waves With Just One Secret Tip PDFDocument4 pagesEasy Way To Spot Elliott Waves With Just One Secret Tip PDFJack XuanNo ratings yet
- Dani Extreme V3 Manual PDFDocument14 pagesDani Extreme V3 Manual PDFHumbertoJaimeNo ratings yet
- Euro CPKDocument20 pagesEuro CPKnovarhai100% (1)
- E702.3M-18 - Acceptance of Concrete Compressive Strength Test Results According To ACI 318M-14Document6 pagesE702.3M-18 - Acceptance of Concrete Compressive Strength Test Results According To ACI 318M-14Christos Leptokaridis100% (1)
- PBL Final ReportDocument4 pagesPBL Final ReportChester Dave Bal-otNo ratings yet
- Síntesis de Procesos: Una Propuesta Holistica Process Synthesis: A Holistic ApproachDocument10 pagesSíntesis de Procesos: Una Propuesta Holistica Process Synthesis: A Holistic ApproachGarcía Nuñez SergioNo ratings yet
- Top Renewable Energy Trends of 2018: Madeleine HoweDocument4 pagesTop Renewable Energy Trends of 2018: Madeleine HowedubryNo ratings yet
- A23.3-04 CorbelsDocument3 pagesA23.3-04 CorbelsDivyesh MistryNo ratings yet
- Crime Scene Power PointDocument31 pagesCrime Scene Power PointAya ChouaibNo ratings yet
- Application of Modern Technology For Fault Diagnosis in Power Transformers Energy ManagementDocument6 pagesApplication of Modern Technology For Fault Diagnosis in Power Transformers Energy ManagementCamila Caceres FloresNo ratings yet
- Wiring DM 1wDocument20 pagesWiring DM 1wAnan NasutionNo ratings yet
- Abraham Tilahun GSE 6950 15Document13 pagesAbraham Tilahun GSE 6950 15Abraham tilahun100% (1)
- Historical FoundationDocument13 pagesHistorical Foundationᜃᜒᜈ᜔ᜎᜒ ᜇᜒᜋᜈ᜔ᜇᜒᜋᜈ᜔No ratings yet
- DMGTDocument139 pagesDMGTPatil TanishkNo ratings yet
- Seminar ReportDocument23 pagesSeminar ReportMamta MishraNo ratings yet
- Heinemann English Language Teaching A Division of Heinemann Publishers (Oxford)Document192 pagesHeinemann English Language Teaching A Division of Heinemann Publishers (Oxford)Tina100% (6)
- III. Performance Task (Photo Analysis)Document14 pagesIII. Performance Task (Photo Analysis)Zayn MendesNo ratings yet
- Short CV RJCDocument3 pagesShort CV RJCShivani KhannaNo ratings yet
- Statistics 1Document149 pagesStatistics 1Zaid AhmedNo ratings yet
- 7 Ways To Better Your LetteringDocument5 pages7 Ways To Better Your LetteringLuciana Freire0% (2)
- Aliquat-336 As A Novel Collector For Quartz FlotationDocument8 pagesAliquat-336 As A Novel Collector For Quartz FlotationMaicol PérezNo ratings yet
- Cantilever Retaining Wall ExampleDocument8 pagesCantilever Retaining Wall ExampleRahul Sinha100% (1)
- Dgms AvaDocument3 pagesDgms AvaVinod GuptaNo ratings yet