You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5796)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (589)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- WORKSHEET 7.2 Gaseous Exchange Across The Respiratory Surfaces and Transport of Gases in HumansDocument5 pagesWORKSHEET 7.2 Gaseous Exchange Across The Respiratory Surfaces and Transport of Gases in HumansFendy Hamid67% (6)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- What Is The Oxygen CascadeDocument27 pagesWhat Is The Oxygen CascadeRandhir Singh90% (10)
- Evaluation of Two Methods To Calculate p50 From A Single Blood SampleDocument3 pagesEvaluation of Two Methods To Calculate p50 From A Single Blood SampleihowardlinNo ratings yet
- CHAPTER 1 - Uses of Biochemical Data - 2014 - Clinical Biochemistry MetabolicDocument5 pagesCHAPTER 1 - Uses of Biochemical Data - 2014 - Clinical Biochemistry Metabolicnour achkarNo ratings yet
- Index 2018 Human-BiochemistryDocument37 pagesIndex 2018 Human-Biochemistrynour achkarNo ratings yet
- Appendix 2 - The Genetic Code - 2018 - Human BiochemistryDocument2 pagesAppendix 2 - The Genetic Code - 2018 - Human Biochemistrynour achkarNo ratings yet
- Index 2014 Clinical-NeuroscienceDocument9 pagesIndex 2014 Clinical-Neurosciencenour achkarNo ratings yet
- List of Abbreviations - 2018 - Human BiochemistryDocument9 pagesList of Abbreviations - 2018 - Human Biochemistrynour achkarNo ratings yet
- CHAPTER 35 - Biochemical Aspects of P - 2014 - Clinical Biochemistry MetabolicDocument10 pagesCHAPTER 35 - Biochemical Aspects of P - 2014 - Clinical Biochemistry Metabolicnour achkarNo ratings yet
- Com. You May Also Complete Your Request On-Line Via The ElsevierDocument1 pageCom. You May Also Complete Your Request On-Line Via The Elseviernour achkarNo ratings yet
- CHAPTER 3 - Quality Aspects of Lab - 2014 - Clinical Biochemistry Metabolic andDocument6 pagesCHAPTER 3 - Quality Aspects of Lab - 2014 - Clinical Biochemistry Metabolic andnour achkarNo ratings yet
- 52 - Hepaciviruses and Hepeviruses Hepatitis C and E Vir - 2012 - Medical MicroDocument9 pages52 - Hepaciviruses and Hepeviruses Hepatitis C and E Vir - 2012 - Medical Micronour achkarNo ratings yet
- Legionella: Legionnaires' Disease Pontiac FeverDocument4 pagesLegionella: Legionnaires' Disease Pontiac Fevernour achkarNo ratings yet
- 39 - Chlamydia Genital and Ocular Infections Infertili - 2012 - Medical MicrobDocument9 pages39 - Chlamydia Genital and Ocular Infections Infertili - 2012 - Medical Microbnour achkarNo ratings yet
- Staphylococcus: Skin Infections Osteomyelitis Bloodstream Infection Food Poisoning Foreign Body Infections MRSADocument7 pagesStaphylococcus: Skin Infections Osteomyelitis Bloodstream Infection Food Poisoning Foreign Body Infections MRSAnour achkarNo ratings yet
- Medical Microbiology: Eighteenth EditionDocument3 pagesMedical Microbiology: Eighteenth Editionnour achkarNo ratings yet
- Index: See Also Peritoneal CavityDocument7 pagesIndex: See Also Peritoneal Cavitynour achkarNo ratings yet
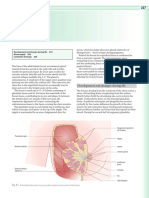
- Chapter 8 - Breast - 2008 - Human AnatomyDocument2 pagesChapter 8 - Breast - 2008 - Human Anatomynour achkarNo ratings yet
- Acknowledgements 2014 Clinical-NeuroscienceDocument1 pageAcknowledgements 2014 Clinical-Neurosciencenour achkarNo ratings yet
- Sources of Electronic Information: WWW - Ncbi.Nlm - Nih.Gov/Pubmed/ WWW - Sgm.Ac - Uk Medbiobrochure/Welcome - HTM WWW - Who.Int/EnDocument1 pageSources of Electronic Information: WWW - Ncbi.Nlm - Nih.Gov/Pubmed/ WWW - Sgm.Ac - Uk Medbiobrochure/Welcome - HTM WWW - Who.Int/Ennour achkarNo ratings yet
- Notices: Working Together To Grow Libraries in Developing CountriesDocument1 pageNotices: Working Together To Grow Libraries in Developing Countriesnour achkarNo ratings yet
- Preface: Neuroanatomy ICT by Crossman and Neary and Has BeenDocument1 pagePreface: Neuroanatomy ICT by Crossman and Neary and Has Beennour achkarNo ratings yet
- Immunity in Bacterial Infections: J. StewartDocument5 pagesImmunity in Bacterial Infections: J. Stewartnour achkarNo ratings yet
- Diagnostic Procedures: R. C. B. SlackDocument8 pagesDiagnostic Procedures: R. C. B. Slacknour achkarNo ratings yet
- Contributors: D. A. A. Ala'Aldeen H. ChartDocument5 pagesContributors: D. A. A. Ala'Aldeen H. Chartnour achkarNo ratings yet
- Preface: From The Preface To The 17 Edition: It Is Now MoreDocument2 pagesPreface: From The Preface To The 17 Edition: It Is Now Morenour achkarNo ratings yet
- Parasitic Infections: Pathogenesis and Immunity: J. StewartDocument7 pagesParasitic Infections: Pathogenesis and Immunity: J. Stewartnour achkarNo ratings yet
- Coryneform Bacteria, Listeria and Erysipelothrix: Diphtheria Listeriosis ErysipeloidDocument12 pagesCoryneform Bacteria, Listeria and Erysipelothrix: Diphtheria Listeriosis Erysipeloidnour achkarNo ratings yet
- Salmonella: Food Poisoning Enteric FeverDocument10 pagesSalmonella: Food Poisoning Enteric Fevernour achkarNo ratings yet
- Immunity in Bacterial Infections: J. StewartDocument5 pagesImmunity in Bacterial Infections: J. Stewartnour achkarNo ratings yet
- The Natural History of Infection: M. R. BarerDocument6 pagesThe Natural History of Infection: M. R. Barernour achkarNo ratings yet
- Streptococcus and Enterococcus: M. KilianDocument16 pagesStreptococcus and Enterococcus: M. Kiliannour achkarNo ratings yet
- Actinomyces, Nocardia and Tropheryma: Actinomycosis Nocardiasis Whipple's DiseaseDocument4 pagesActinomyces, Nocardia and Tropheryma: Actinomycosis Nocardiasis Whipple's Diseasenour achkarNo ratings yet
- BloodDocument27 pagesBloodRagda AhmedNo ratings yet
- Praktikum BiokimiaDocument20 pagesPraktikum BiokimiaEndah Sekar PalupiNo ratings yet
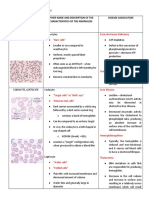
- RBC AnomaliesDocument10 pagesRBC AnomaliesSHUPATUSSAINo ratings yet
- Biochemistry Quizzes by Ronnie BaticulonDocument20 pagesBiochemistry Quizzes by Ronnie BaticulonMhartin GarciaNo ratings yet
- Report HB ELECTROPHORESISDocument2 pagesReport HB ELECTROPHORESISShweta PatilNo ratings yet
- OET30 Reading Day 11 - 20Document124 pagesOET30 Reading Day 11 - 20Shraadha JaiswaniNo ratings yet
- GCE Biology-Human Biology-Teachers' Guide Revised 18-02-14 PDFDocument93 pagesGCE Biology-Human Biology-Teachers' Guide Revised 18-02-14 PDFTara MillsNo ratings yet
- 2507 CAP3 HB - enDocument29 pages2507 CAP3 HB - enTrường Bửu TrầnNo ratings yet
- Tissue OpticsDocument55 pagesTissue OpticsNajihah AnisNo ratings yet
- 02Document12 pages02nayan159No ratings yet
- Physiology Summary Chapter 32Document6 pagesPhysiology Summary Chapter 32gail018No ratings yet
- SGD Case: Hematology Weakness: Dizziness: Anorexia: Vomit: Numbness: TinglingingDocument10 pagesSGD Case: Hematology Weakness: Dizziness: Anorexia: Vomit: Numbness: TinglingingEveryday FundayNo ratings yet
- Text Book On Practical Kriya Sharira PDFDocument13 pagesText Book On Practical Kriya Sharira PDFCHAUKHAMBHA PRAKASHAK50% (6)
- 4 HypoxiaDocument37 pages4 HypoxiaPauloNo ratings yet
- Crocodile Haemoglobin Article QuestionsDocument1 pageCrocodile Haemoglobin Article Questionsapi-238643412No ratings yet
- 75 FullDocument8 pages75 FullFede0No ratings yet
- Pulse Oximetry - UpToDateDocument23 pagesPulse Oximetry - UpToDateHardian RakhmawardanaNo ratings yet
- Normal Lab Values UsmleDocument3 pagesNormal Lab Values UsmleAnne Maria50% (2)
- Dr. Namrata .A (B.A.M.S - Mumbai)Document23 pagesDr. Namrata .A (B.A.M.S - Mumbai)kalloornatorNo ratings yet
- Case Presentation - OsteomyelitisDocument58 pagesCase Presentation - Osteomyelitisbernadette_olaveNo ratings yet
- Chemistry Practice Olympiad Stage 1Document27 pagesChemistry Practice Olympiad Stage 1DevYShethNo ratings yet
- 22 Respiratory SystemDocument99 pages22 Respiratory SystemvanderphysNo ratings yet
- 14 Diagnostic Value of HematologyDocument24 pages14 Diagnostic Value of HematologyPablo LópezNo ratings yet
- Subjective Cues: Ineffective Tissue PerfusionDocument3 pagesSubjective Cues: Ineffective Tissue PerfusionFlorence Lacea Rotsap-DeguitNo ratings yet
- Iron Deficiency Anemia-Pathophysiology and Lab DiagnosisDocument69 pagesIron Deficiency Anemia-Pathophysiology and Lab Diagnosisswathi bsNo ratings yet
- HL Paper3Document28 pagesHL Paper3Yuvraj GuptaNo ratings yet
- HealthDocument319 pagesHealthpredic1No ratings yet