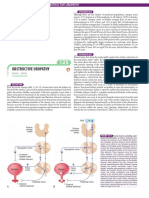
You might also like
- YASHU Final Technical Seminar ReportDocument23 pagesYASHU Final Technical Seminar Reportyashwanth vNo ratings yet
- Vistara Jan 2022Document43 pagesVistara Jan 2022Dheeraj GaurNo ratings yet
- The Impact of Indias Cyber Security Law and Cyber Forensic On Building Techno-Centric Smartcity IoT EnvironmentDocument9 pagesThe Impact of Indias Cyber Security Law and Cyber Forensic On Building Techno-Centric Smartcity IoT EnvironmentNandini NanduNo ratings yet
- Advanced Real-Time Systems ARTIST Project IST-2001-34820 BMW 2004Document372 pagesAdvanced Real-Time Systems ARTIST Project IST-2001-34820 BMW 2004كورسات هندسيةNo ratings yet
- 15 10 2021 InglesDocument3 pages15 10 2021 InglesJose DavidNo ratings yet
- GBG Idscan Ieos Web API v4Document16 pagesGBG Idscan Ieos Web API v4Laberto KelenNo ratings yet
- Program Book Rev 8Document115 pagesProgram Book Rev 8TuswanNo ratings yet
- Small Scale Wind Tunnel Testing of Model PropellersDocument10 pagesSmall Scale Wind Tunnel Testing of Model PropellersClaus WehmannNo ratings yet
- DSTL - Lab ManualDocument28 pagesDSTL - Lab ManualGud GuyNo ratings yet
- Analysis of E-Waste Recycling Behavior Based On Survey at A Midwestern PDFDocument9 pagesAnalysis of E-Waste Recycling Behavior Based On Survey at A Midwestern PDFSandamini KaushalyaNo ratings yet
- Load Balancing in Cloud ComputingDocument14 pagesLoad Balancing in Cloud ComputingAnonymous CwJeBCAXpNo ratings yet
- Data Sheet: SAW DuplexerDocument24 pagesData Sheet: SAW DuplexerHongdang DinhNo ratings yet
- IOT Project POSTERDocument1 pageIOT Project POSTERFemilNo ratings yet
- Virus Explorer-WorksheetDocument5 pagesVirus Explorer-WorksheetDiane Princess SultanNo ratings yet
- Agastya: HPC User ManualDocument73 pagesAgastya: HPC User ManualromNo ratings yet
- AR CyberDocument6 pagesAR Cyberchernet kebedeNo ratings yet
- Akshay Bora HifieldDocument62 pagesAkshay Bora Hifieldsamadhan landgeNo ratings yet
- Chapter 1 Gec 5Document17 pagesChapter 1 Gec 5Mabini BernadaNo ratings yet
- Quantum Information and Computation - NatureDocument10 pagesQuantum Information and Computation - Nature*83*22*No ratings yet
- ALTTC Project Report FullDocument24 pagesALTTC Project Report FullKrishna ShahNo ratings yet
- The Great-Resignation - TimeDocument3 pagesThe Great-Resignation - TimeCláudia AntunesNo ratings yet
- PelangiDocument13 pagesPelangiFAHMA IQBALNo ratings yet
- VX Series User ManualDocument121 pagesVX Series User Manualthangpv94No ratings yet
- Delta 1kW Wireless Charging System AppNote en V01 20200922Document38 pagesDelta 1kW Wireless Charging System AppNote en V01 20200922UUasin KaowNo ratings yet
- An Introductory Guide To Computational Methods For The Solution of Physics ProblemsDocument227 pagesAn Introductory Guide To Computational Methods For The Solution of Physics ProblemsCesar SalasNo ratings yet
- Pulsar 160 Ns Users GuideDocument2 pagesPulsar 160 Ns Users GuideaunerNo ratings yet
- Lectures On Computational Fluid Dynamics PDFDocument205 pagesLectures On Computational Fluid Dynamics PDFfsnexussNo ratings yet
- Po KCenDocument413 pagesPo KCenBorko SimicNo ratings yet
- 80 Python Interview Questions & AnswersDocument23 pages80 Python Interview Questions & AnswersMahmoud ElhadyNo ratings yet
- A Comprehensive Review of Wireless Body Area Network - 2019 - A Comprehensive Review of Wireless Body Area NetworkDocument1 pageA Comprehensive Review of Wireless Body Area Network - 2019 - A Comprehensive Review of Wireless Body Area NetworkGeneration GenerationNo ratings yet
- Bài tập tự động hóa sản xuất - 0001-0001Document1 pageBài tập tự động hóa sản xuất - 0001-0001Minh NhậtNo ratings yet
- UPS User ManualDocument52 pagesUPS User ManualStephen Rey CaldeaNo ratings yet
- EC-Council Certified Incident Handler (ECIH) Course SummaryDocument7 pagesEC-Council Certified Incident Handler (ECIH) Course SummaryVinay KumarNo ratings yet
- ENKOnail Nail MachineDocument1 pageENKOnail Nail MachinelipezaoNo ratings yet
- (Cambridge Monographs on Applied and Computational Mathematics) Alfio Quarteroni, Luca Dede', Andrea Manzoni, Christian Vergara - Mathematical Modelling of the Human Cardiovascular System_ Data, NumerDocument294 pages(Cambridge Monographs on Applied and Computational Mathematics) Alfio Quarteroni, Luca Dede', Andrea Manzoni, Christian Vergara - Mathematical Modelling of the Human Cardiovascular System_ Data, NumerNarendra DesaiNo ratings yet
- Face Spoofing Attacks and Detection: A ReviewDocument5 pagesFace Spoofing Attacks and Detection: A ReviewOm DandadeNo ratings yet
- Specifications: KID Teer OaderDocument2 pagesSpecifications: KID Teer OaderArvind HarryNo ratings yet
- $$TR Avt 275 AllDocument260 pages$$TR Avt 275 AllFendy FendersonNo ratings yet
- Kelompok 1. Object Detection in A Maritime Environment - CompressedDocument16 pagesKelompok 1. Object Detection in A Maritime Environment - CompressedRumah sakitNo ratings yet
- Coarse Graining Shannon and Von Neumann EntropiesDocument26 pagesCoarse Graining Shannon and Von Neumann Entropiesdani_cirigNo ratings yet
- Disney PatentDocument12 pagesDisney PatentCharles GrossNo ratings yet
- S-3109 FinalDocument60 pagesS-3109 FinalRaúl Oscar LedesmaNo ratings yet
- Characterization Techniques For NanoparticlesDocument8 pagesCharacterization Techniques For NanoparticlesRichard J. GrayNo ratings yet
- SUN2000-50KTL-M3 EN50549-1 Cert - Intertek - 20221018Document2 pagesSUN2000-50KTL-M3 EN50549-1 Cert - Intertek - 20221018Stefan BusoiNo ratings yet
- As 129959 Iv3-G500ca Iv3-G500ma Iv3-G600ca Im 96M17460 WW GB 2102 1Document2 pagesAs 129959 Iv3-G500ca Iv3-G500ma Iv3-G600ca Im 96M17460 WW GB 2102 1AfifNasirNo ratings yet
- Periodic Modulation and Functional Demonstrations of Mechanically Operated Reciprocating VentilatorDocument8 pagesPeriodic Modulation and Functional Demonstrations of Mechanically Operated Reciprocating VentilatorIJRASETPublications100% (1)
- Bpo ProcessDocument4 pagesBpo ProcessAvick ChakrabortyNo ratings yet
- Review of Laser Doping and Its Applications in Silicon Solar CellsDocument12 pagesReview of Laser Doping and Its Applications in Silicon Solar CellsJIJIN KNo ratings yet
- Cs230exam Spr18 Soln PDFDocument45 pagesCs230exam Spr18 Soln PDFMOHAMMAD100% (1)
- CABPD ManualDocument35 pagesCABPD ManualSupritha KNo ratings yet
- 978 3 642 32784 1Document266 pages978 3 642 32784 1pepe pepitosNo ratings yet
- Deep Learning in Mining Biological DataDocument33 pagesDeep Learning in Mining Biological DataCarlos Ramos100% (1)
- School of Education - Scholarly WritingDocument3 pagesSchool of Education - Scholarly WritingJohn M. NdunguNo ratings yet
- W1 - Lecture - Vectors - Part 1 (Updated)Document39 pagesW1 - Lecture - Vectors - Part 1 (Updated)Nam ĐinhNo ratings yet
- Vastipatrak 17oct 10Document18 pagesVastipatrak 17oct 10sonaldivNo ratings yet
- A. 100 B. BTWN 25 and 26 Value: (0+1) /2 0.5 BTWN 75 and 76 Value: (4+4) /2 4 CDocument3 pagesA. 100 B. BTWN 25 and 26 Value: (0+1) /2 0.5 BTWN 75 and 76 Value: (4+4) /2 4 CAruzhan ZhadyrassynNo ratings yet
- 1 Photoacoustic Signal Generation For Underwater Communication by Ayele Ossa LekaDocument45 pages1 Photoacoustic Signal Generation For Underwater Communication by Ayele Ossa Lekaayele ossaNo ratings yet
- Using Isometries For Computational Design and FabricationDocument12 pagesUsing Isometries For Computational Design and FabricationCaigui JiangNo ratings yet
- 1 s2.0 S0029801822002840 MainDocument13 pages1 s2.0 S0029801822002840 Main刘五六No ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument41 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsArya ChowdhuryNo ratings yet
- CAKIR-YALCIN-ATES Tarım Makineleri Dergisi 2009Document5 pagesCAKIR-YALCIN-ATES Tarım Makineleri Dergisi 2009fadimeatesNo ratings yet
- OZKAN-ATES-KARA-TURAN GAMBOA - TAMMETIN Horticulturae 2022Document14 pagesOZKAN-ATES-KARA-TURAN GAMBOA - TAMMETIN Horticulturae 2022fadimeatesNo ratings yet
- 2023 Atif Ates Unal Takma AltindisliDocument24 pages2023 Atif Ates Unal Takma AltindislifadimeatesNo ratings yet
- Cetinkaya-Ates J.turk Phytopath-2016-2 PDFDocument15 pagesCetinkaya-Ates J.turk Phytopath-2016-2 PDFfadimeatesNo ratings yet
- Cetinkaya-Ates The J of Turkish Phytopathology 45 (2-3) 2016-Tam MetinDocument8 pagesCetinkaya-Ates The J of Turkish Phytopathology 45 (2-3) 2016-Tam MetinfadimeatesNo ratings yet
- Okur Kayikcioglu Ates Yagmur 2016Document14 pagesOkur Kayikcioglu Ates Yagmur 2016fadimeatesNo ratings yet
- KESKIN-KAYA-ATES-TURAN-GAMBOA-Plants 2022Document12 pagesKESKIN-KAYA-ATES-TURAN-GAMBOA-Plants 2022fadimeatesNo ratings yet
- Ates-Coban - Kara-Sabir-J. Bulgaria-2011 PDFDocument11 pagesAtes-Coban - Kara-Sabir-J. Bulgaria-2011 PDFfadimeatesNo ratings yet
- Karakus Ates Keskin Turan Kaya 2023 Mitteilungen KlosterneuburgDocument16 pagesKarakus Ates Keskin Turan Kaya 2023 Mitteilungen KlosterneuburgfadimeatesNo ratings yet
- KAYA-INCESU-ATES-KESKIN-VERDUGO-VAQUEZ-GAMBOA-Plants 2022Document14 pagesKAYA-INCESU-ATES-KESKIN-VERDUGO-VAQUEZ-GAMBOA-Plants 2022fadimeatesNo ratings yet
- ATES-KAYA 2021 - Article - TheRelationshipBetweenIronAndNDocument7 pagesATES-KAYA 2021 - Article - TheRelationshipBetweenIronAndNfadimeatesNo ratings yet
- ATES-Erwerbs-Obstbau 2022Document6 pagesATES-Erwerbs-Obstbau 2022fadimeatesNo ratings yet
- Incesu Karakus Seyed Hajizadeh, Ates.. Plants 2022 Tam MetinDocument11 pagesIncesu Karakus Seyed Hajizadeh, Ates.. Plants 2022 Tam MetinfadimeatesNo ratings yet
- OZKAN-ATES-KARA-TURAN GAMBOA - TAMMETIN Horticulturae 2022Document14 pagesOZKAN-ATES-KARA-TURAN GAMBOA - TAMMETIN Horticulturae 2022fadimeatesNo ratings yet
- ATES-Erwerbs-Obstbau 2022Document6 pagesATES-Erwerbs-Obstbau 2022fadimeatesNo ratings yet
- Emerald-Crown - DOMINOES PDFDocument25 pagesEmerald-Crown - DOMINOES PDFEC Pisano Riggio70% (20)
- KAYA-INCESU-ATES-KESKIN-VERDUGO-VAQUEZ-GAMBOA-Plants 2022Document14 pagesKAYA-INCESU-ATES-KESKIN-VERDUGO-VAQUEZ-GAMBOA-Plants 2022fadimeatesNo ratings yet
- The Little Match GirlDocument39 pagesThe Little Match GirlEvelyn Andrea Parra Arellano90% (10)
- KESKIN-KAYA-ATES-TURAN-GAMBOA-Plants 2022Document12 pagesKESKIN-KAYA-ATES-TURAN-GAMBOA-Plants 2022fadimeatesNo ratings yet
- (Discovering Holistic Health) Stefan Ball - Principles of Bach Flower Remedies - What It Is, How It Works, and What It Can Do For You-Jessica Kingsley Publishers (2013) PDFDocument128 pages(Discovering Holistic Health) Stefan Ball - Principles of Bach Flower Remedies - What It Is, How It Works, and What It Can Do For You-Jessica Kingsley Publishers (2013) PDFfadimeates100% (1)
- CinderellaDocument9 pagesCinderellaGolpo guccoNo ratings yet
- Sinbad Oxford Dominoes Starter PDFDocument57 pagesSinbad Oxford Dominoes Starter PDFLuz Castellanos92% (13)
- Vitamin and Mineral CardsDocument11 pagesVitamin and Mineral CardsfadimeatesNo ratings yet
- (Read It Yourself 1) - Goldilocks and The Three Bears - Pearson PLC, Penguin Group, Ladybird Books (1998)Document15 pages(Read It Yourself 1) - Goldilocks and The Three Bears - Pearson PLC, Penguin Group, Ladybird Books (1998)fadimeatesNo ratings yet
- Oscar Wilde - The Happy Prince (1995)Document149 pagesOscar Wilde - The Happy Prince (1995)fadimeatesNo ratings yet
- Model of Vines Production Variation in Relation To Physiological IndicesDocument10 pagesModel of Vines Production Variation in Relation To Physiological IndicesfadimeatesNo ratings yet
- Aunt C Hemochromatosis Sanf 16 THis No Date SanfDocument18 pagesAunt C Hemochromatosis Sanf 16 THis No Date SanffadimeatesNo ratings yet
- Raisins in Human Health: A Review: 39 World Congress of Vine and WineDocument6 pagesRaisins in Human Health: A Review: 39 World Congress of Vine and WinefadimeatesNo ratings yet
- Agnes Fay Moegan, Louise Kimmel, Anna Field AND Paul F. NicholsDocument14 pagesAgnes Fay Moegan, Louise Kimmel, Anna Field AND Paul F. NicholsfadimeatesNo ratings yet
- Ajph 25 3 328Document8 pagesAjph 25 3 328fadimeatesNo ratings yet
- Abdomen and Superficial Structures (Diagnostic Medical Sonography Series) Fourth EditionDocument1,232 pagesAbdomen and Superficial Structures (Diagnostic Medical Sonography Series) Fourth EditionBenito Trevizo100% (2)
- OU SumberDocument5 pagesOU SumberAnggaNo ratings yet
- Localisation of Brain FunctionDocument1 pageLocalisation of Brain FunctionaryanNo ratings yet
- Lesson 4 Circulatory System 2Document2 pagesLesson 4 Circulatory System 2Glaisa CudiaNo ratings yet
- Postoperative Nausea and VomitingDocument23 pagesPostoperative Nausea and Vomitingsuleman2009No ratings yet
- The Neurology of Acquired Pedophilia: NeurocaseDocument13 pagesThe Neurology of Acquired Pedophilia: NeurocaseMaria Isabel Montañez RestrepoNo ratings yet
- Sih Ar 2015Document230 pagesSih Ar 2015Ali SaeedNo ratings yet
- A Comprehensive Review of The Clitoris and Its Role in FemaleDocument19 pagesA Comprehensive Review of The Clitoris and Its Role in FemaleIsis StoreNo ratings yet
- Fluid Flow and Mass Transport in Brain TissueDocument33 pagesFluid Flow and Mass Transport in Brain TissueMango SteenNo ratings yet
- Danger Space - WikipediaDocument7 pagesDanger Space - WikipediaMedhat SabonNo ratings yet
- Basic Terminologies in Dental Anatomy: Group 01Document39 pagesBasic Terminologies in Dental Anatomy: Group 01Thiên Lượng100% (1)
- Bilateral Vestibulopathy: Diagnostic CriteriaDocument22 pagesBilateral Vestibulopathy: Diagnostic CriteriaHerminaElenaNo ratings yet
- Analysis of Head CT Scans Flagged by Deep Learning Software For Acute Intracranial HemorrhageDocument6 pagesAnalysis of Head CT Scans Flagged by Deep Learning Software For Acute Intracranial HemorrhageAshish MehtaNo ratings yet
- Skeletal System WorksheetDocument7 pagesSkeletal System Worksheet마스러버No ratings yet
- Metameric ManualDocument160 pagesMetameric ManualRizal Ting100% (2)
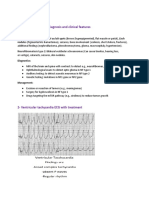
- Unit 6 - Topic 6 - Human Physiologyy - (11 HL and SL)Document13 pagesUnit 6 - Topic 6 - Human Physiologyy - (11 HL and SL)John DoeNo ratings yet
- Mandolesi Et Al, 2018Document11 pagesMandolesi Et Al, 2018Mateus CarvalhoNo ratings yet
- Comparison of CT and MR Imaging in Ischemic StrokeDocument10 pagesComparison of CT and MR Imaging in Ischemic Strokeتغريد عبادةNo ratings yet
- Parotidectomy: H.Shameer AhamedDocument47 pagesParotidectomy: H.Shameer AhamedAndreas RendraNo ratings yet
- Eye Diagnostic PointsDocument22 pagesEye Diagnostic PointsShoaib AkhtarNo ratings yet
- 2nd Pharmacology Sessional PaperDocument2 pages2nd Pharmacology Sessional PaperRaman NaiknawareNo ratings yet
- Sciatica Nerve (Causes and Treatment)Document4 pagesSciatica Nerve (Causes and Treatment)esmollahanNo ratings yet
- Medicine OSPE Pre AnnualDocument17 pagesMedicine OSPE Pre AnnualMunawarNo ratings yet
- Liver HistologyDocument7 pagesLiver HistologyAhmad Khan GhoriNo ratings yet
- Guide To The Assessment of The Degree of Permanent Impairment The Guide Edition 2.1 of The Guide PDF, 1.84 MBDocument332 pagesGuide To The Assessment of The Degree of Permanent Impairment The Guide Edition 2.1 of The Guide PDF, 1.84 MBbhaskarjalanNo ratings yet
- ANPS 020 Cornbrooks 02-07-14Document25 pagesANPS 020 Cornbrooks 02-07-14enam professorNo ratings yet
- A. I. Ii. Iii.: Protection. Serving As The Covering of The Body, ItDocument9 pagesA. I. Ii. Iii.: Protection. Serving As The Covering of The Body, ItFrancis100% (1)
- Learning Objectives:: at The End of This Lecture, You Will Be Able ToDocument119 pagesLearning Objectives:: at The End of This Lecture, You Will Be Able ToMaica LectanaNo ratings yet
- Final Hydrocephalus Care PlanDocument11 pagesFinal Hydrocephalus Care PlanSAYMABANUNo ratings yet
- Dho Health Science Updated 8th Edition Simmers Test BankDocument9 pagesDho Health Science Updated 8th Edition Simmers Test Bankjosephrodriguez04121993gbm100% (27)
- The Age of Magical Overthinking: Notes on Modern IrrationalityFrom EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityRating: 4 out of 5 stars4/5 (23)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisFrom EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisRating: 4.5 out of 5 stars4.5/5 (42)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsFrom EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNo ratings yet
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedRating: 5 out of 5 stars5/5 (80)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsFrom EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsRating: 5 out of 5 stars5/5 (1)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaFrom EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaRating: 4.5 out of 5 stars4.5/5 (266)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeFrom EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeNo ratings yet
- The Obesity Code: Unlocking the Secrets of Weight LossFrom EverandThe Obesity Code: Unlocking the Secrets of Weight LossRating: 4 out of 5 stars4/5 (5)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Sleep Stories for Adults: Overcome Insomnia and Find a Peaceful AwakeningFrom EverandSleep Stories for Adults: Overcome Insomnia and Find a Peaceful AwakeningRating: 4 out of 5 stars4/5 (3)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.From EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Rating: 4.5 out of 5 stars4.5/5 (110)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsFrom EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsRating: 3.5 out of 5 stars3.5/5 (3)
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDFrom EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDRating: 5 out of 5 stars5/5 (1)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- An Autobiography of Trauma: A Healing JourneyFrom EverandAn Autobiography of Trauma: A Healing JourneyRating: 5 out of 5 stars5/5 (2)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsFrom EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsRating: 4.5 out of 5 stars4.5/5 (169)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessFrom EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessRating: 4.5 out of 5 stars4.5/5 (328)
- Outlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisFrom EverandOutlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisRating: 4 out of 5 stars4/5 (1)
- Summary: The Myth of Normal: Trauma, Illness, and Healing in a Toxic Culture By Gabor Maté MD & Daniel Maté: Key Takeaways, Summary & AnalysisFrom EverandSummary: The Myth of Normal: Trauma, Illness, and Healing in a Toxic Culture By Gabor Maté MD & Daniel Maté: Key Takeaways, Summary & AnalysisRating: 4 out of 5 stars4/5 (9)
- 12 Rules for Life by Jordan B. Peterson - Book Summary: An Antidote to ChaosFrom Everand12 Rules for Life by Jordan B. Peterson - Book Summary: An Antidote to ChaosRating: 4.5 out of 5 stars4.5/5 (207)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryFrom EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryRating: 4 out of 5 stars4/5 (44)
- The Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeFrom EverandThe Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeRating: 4.5 out of 5 stars4.5/5 (253)