You might also like
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Liu Et Al-2013-Journal of Pineal ResearchDocument10 pagesLiu Et Al-2013-Journal of Pineal ResearchElena Martínez CelisNo ratings yet
- Naskar Et Al-2013-Journal of Pineal ResearchDocument9 pagesNaskar Et Al-2013-Journal of Pineal ResearchElena Martínez CelisNo ratings yet
- Lima-Cabello Et Al-2014-Journal of Pineal ResearchDocument9 pagesLima-Cabello Et Al-2014-Journal of Pineal ResearchElena Martínez CelisNo ratings yet
- Juan Et Al-2014-Journal of Pineal ResearchDocument11 pagesJuan Et Al-2014-Journal of Pineal ResearchElena Martínez CelisNo ratings yet
- Kaur Et Al-2010-Journal of Pineal ResearchDocument9 pagesKaur Et Al-2010-Journal of Pineal ResearchElena Martínez CelisNo ratings yet
- Lacoste Et Al-2015-Journal of Pineal ResearchDocument21 pagesLacoste Et Al-2015-Journal of Pineal ResearchElena Martínez CelisNo ratings yet
- The Microtubule Cytoskeleton Acts As A Key Downstream Effector of Neurotransmitter SignalingDocument8 pagesThe Microtubule Cytoskeleton Acts As A Key Downstream Effector of Neurotransmitter SignalingElena Martínez CelisNo ratings yet
- MELATONINA F ACTINA Matsui - Et - Al-1997-Journal - of - Pineal - ResearchDocument7 pagesMELATONINA F ACTINA Matsui - Et - Al-1997-Journal - of - Pineal - ResearchElena Martínez CelisNo ratings yet
- Tau Filaments in Neurodegenerative Diseases Goedert-2018-FEBS - LettersDocument30 pagesTau Filaments in Neurodegenerative Diseases Goedert-2018-FEBS - LettersElena Martínez CelisNo ratings yet
- G-Actina Espinas DendriticasDocument3 pagesG-Actina Espinas DendriticasElena Martínez CelisNo ratings yet
- Phosporilation Tau EsquemaDocument9 pagesPhosporilation Tau EsquemaElena Martínez CelisNo ratings yet
- Actin-Based Plasticity in Dendritic Spines: References and NotesDocument6 pagesActin-Based Plasticity in Dendritic Spines: References and NotesElena Martínez CelisNo ratings yet
- Calcium in SpinesDocument6 pagesCalcium in SpinesElena Martínez CelisNo ratings yet
- Radiographic Positioning DogDocument1 pageRadiographic Positioning DogElena Martínez Celis100% (2)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Chap 4 Section 3 Review Q&A'sDocument3 pagesChap 4 Section 3 Review Q&A'sAashi PatelNo ratings yet
- BIOL 215 - CWRU Final Exam Learning GoalsDocument32 pagesBIOL 215 - CWRU Final Exam Learning GoalsKesharaSSNo ratings yet
- Tatva CellDocument66 pagesTatva CellGuriya Kumari100% (1)
- Cell, Tissue & MembraneDocument24 pagesCell, Tissue & MembranehumaNo ratings yet
- The Periodontal Ligament: A Unique, Multifunctional Connective TissueDocument21 pagesThe Periodontal Ligament: A Unique, Multifunctional Connective TissueSamuel Flores CalderonNo ratings yet
- 1 4927010714729578637Document368 pages1 4927010714729578637siddhantaNo ratings yet
- Unit I Module 2: Prokaryotic and Eukaryotic CellsDocument8 pagesUnit I Module 2: Prokaryotic and Eukaryotic CellsMirunalini GobinathNo ratings yet
- Cell Locomotion PDFDocument8 pagesCell Locomotion PDFmanoj_rkl_07No ratings yet
- Chapter 3 The Cellular Level of Organization: Fundamentals of Anatomy and Physiology, 11e (Martini)Document50 pagesChapter 3 The Cellular Level of Organization: Fundamentals of Anatomy and Physiology, 11e (Martini)RO OMNo ratings yet
- Scanning Ion Microscopy and Its Application in MicrobiologyDocument9 pagesScanning Ion Microscopy and Its Application in MicrobiologyAthenaeum Scientific PublishersNo ratings yet
- Cells Exploration ActivitiesDocument16 pagesCells Exploration ActivitiesLouiseNo ratings yet
- Cell Nanomaterial InteractionsDocument42 pagesCell Nanomaterial Interactionslux0008No ratings yet
- QB Biology Chapter 5Document19 pagesQB Biology Chapter 5Holda MohammedaliNo ratings yet
- Veterinary Histology 1532547893Document233 pagesVeterinary Histology 1532547893Teguh ArinataNo ratings yet
- Final Syllabi - Microbiology 10062019Document57 pagesFinal Syllabi - Microbiology 10062019JharkhandNo ratings yet
- 6th Science PDE Worksheet 2018 1 TermDocument5 pages6th Science PDE Worksheet 2018 1 TermJUAN CARLOS0% (1)
- Capitulo 1 Webster and Weber Introduction To FungiDocument39 pagesCapitulo 1 Webster and Weber Introduction To Fungidianacarranza148No ratings yet
- Cell-Structure and FunctionDocument11 pagesCell-Structure and Functionانس خيرناNo ratings yet
- Biomolecules and Cells NotesDocument81 pagesBiomolecules and Cells NotesLily RobinsonNo ratings yet
- Biochemistry of LensDocument24 pagesBiochemistry of LensriskhapangestikaNo ratings yet
- Color and Label Color and Label .: Cells and Their OrganellesDocument4 pagesColor and Label Color and Label .: Cells and Their OrganellesEllie GriffinNo ratings yet
- Chapter 6 Part 11Document2 pagesChapter 6 Part 11dandymontezNo ratings yet
- Using Cell Structures To Develop Functional Nanomaterials and Nanostructures-Case Studies of Actin Filaments and MicrotubulesDocument11 pagesUsing Cell Structures To Develop Functional Nanomaterials and Nanostructures-Case Studies of Actin Filaments and Microtubules健康新體驗No ratings yet
- Cell Structure and FunctionDocument2 pagesCell Structure and FunctionSynde Galinato DomaoanNo ratings yet
- Cytoplasm and Cytoskelton Practice WorksheetDocument4 pagesCytoplasm and Cytoskelton Practice WorksheetMKGU100% (1)
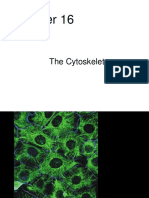
- Chapter16 The CytoskeletonDocument83 pagesChapter16 The CytoskeletonRonNo ratings yet
- CytoskeletonDocument2 pagesCytoskeletonastro paculanNo ratings yet
- MSC MPlil ZoologyDocument18 pagesMSC MPlil ZoologyYougesh KumarNo ratings yet
- Biochemistry PDFDocument431 pagesBiochemistry PDFNimhans1234589% (18)
- 2 MODULE CHAPTER 3&4 AnaPhy 2020-2021 PDFDocument51 pages2 MODULE CHAPTER 3&4 AnaPhy 2020-2021 PDFJosh MagatNo ratings yet