You might also like
- Endemic Species of KoreaDocument401 pagesEndemic Species of KoreaentoNo ratings yet
- Plant Systematics by Op Sharma Chapter 01Document9 pagesPlant Systematics by Op Sharma Chapter 01MUHAMMAD MUBASHIR ALINo ratings yet
- Theories of Evolution WorksheetDocument3 pagesTheories of Evolution WorksheetManroop NijjarNo ratings yet
- Darwin Vs Lamarck Worksheet PDFDocument2 pagesDarwin Vs Lamarck Worksheet PDFZhiyong Huang100% (2)
- Desert Biology: Special Topics on the Physical and Biological Aspects of Arid RegionsFrom EverandDesert Biology: Special Topics on the Physical and Biological Aspects of Arid RegionsG. W. BrownNo ratings yet
- 2010 - Book - Soil Biology and Agriculture in The TropicsDocument330 pages2010 - Book - Soil Biology and Agriculture in The TropicsKarol Gonzalez100% (1)
- Chapter 2-1 Protozoa Diversity: Moss-Dwelling Micro-OrganismsDocument16 pagesChapter 2-1 Protozoa Diversity: Moss-Dwelling Micro-OrganismsJose Facundo Ortega OrizNo ratings yet
- S204 Book 1 - The Open UniversityDocument144 pagesS204 Book 1 - The Open UniversitySara Cooper100% (1)
- May (1994)Document8 pagesMay (1994)star warsNo ratings yet
- Tropical Rainforest and Tropical GrasslandDocument7 pagesTropical Rainforest and Tropical GrasslandGellie Dela Rosa ValenciaNo ratings yet
- Antonelli - Etal.2018.amazonina DiversityDocument6 pagesAntonelli - Etal.2018.amazonina Diversitycamila zapataNo ratings yet
- Mortality Polinices (B.T)Document8 pagesMortality Polinices (B.T)Nadia ZaharaNo ratings yet
- Bahasa Inggris GeografiDocument11 pagesBahasa Inggris GeografiElia RizkiaNo ratings yet
- Amazonia Is The Primary Source of Neotropical BiodiversityDocument6 pagesAmazonia Is The Primary Source of Neotropical BiodiversitySamuel FreitasNo ratings yet
- Society For The Study of EvolutionDocument9 pagesSociety For The Study of EvolutionWenner BritoNo ratings yet
- 1950 - Thorson - Larval EcologyDocument45 pages1950 - Thorson - Larval Ecologyetejada00No ratings yet
- Species Richness in Tropical Forests AshtonDocument7 pagesSpecies Richness in Tropical Forests Ashtonfrancisco2and2aNo ratings yet
- 8.polis 1999Document13 pages8.polis 1999MarioLowellNo ratings yet
- Diversity of The Families of Coleoptera Captured With Pitfall Traps in Five Different Environments in Santa Maria, RS, BrazilDocument10 pagesDiversity of The Families of Coleoptera Captured With Pitfall Traps in Five Different Environments in Santa Maria, RS, BrazilSarai Olmedo CruzNo ratings yet
- Polychaete zonation patterns in Santos estuaryDocument10 pagesPolychaete zonation patterns in Santos estuaryManuelle da SilvaNo ratings yet
- 36 ch13Document23 pages36 ch13PraveenNo ratings yet
- Polinizacion ChontaduroDocument14 pagesPolinizacion ChontaduroDiianis CortesNo ratings yet
- Half Fter 2002Document11 pagesHalf Fter 2002Jonathan PerezNo ratings yet
- Ieltstest 1Document9 pagesIeltstest 1Bùi Hoàng DũngNo ratings yet
- 13-Organisms and Populations (Extra Questions)Document32 pages13-Organisms and Populations (Extra Questions)Poonam PathakNo ratings yet
- Costello, May, Stork - 2013 - Can We Name Earth ' S Species Before They Go ExtinctDocument5 pagesCostello, May, Stork - 2013 - Can We Name Earth ' S Species Before They Go ExtinctImanol O'ConnorNo ratings yet
- Why We Care About Soil FaunaDocument4 pagesWhy We Care About Soil FaunaAnjusNo ratings yet
- 5-Costello 2017 Marine Biogeographic Realms and Species EndemicityDocument10 pages5-Costello 2017 Marine Biogeographic Realms and Species EndemicityLeah Marie MahomocNo ratings yet
- Paper Springer Klemp 16 06Document24 pagesPaper Springer Klemp 16 06Finn KlempNo ratings yet
- Envi-Bio LecDocument15 pagesEnvi-Bio LecIslam SamirNo ratings yet
- Chapter 1 - IntroductionDocument10 pagesChapter 1 - IntroductionGilson CarlosNo ratings yet
- Biotic and Abiotic Factors Shaping Ant (Hymenoptera: Formicidae) Assemblages in Brazilian Coastal Sand Dunes: The Case of Restinga in Santa CatarinaDocument11 pagesBiotic and Abiotic Factors Shaping Ant (Hymenoptera: Formicidae) Assemblages in Brazilian Coastal Sand Dunes: The Case of Restinga in Santa CatarinaJustine PamaNo ratings yet
- J Vegetation Science - 2012 - Wilson - Plant Species Richness The World RecordsDocument7 pagesJ Vegetation Science - 2012 - Wilson - Plant Species Richness The World RecordsAhmad AlawiNo ratings yet
- ArtigoDocument9 pagesArtigoCylene CâmaraNo ratings yet
- Nature Vol. 259 January 29 1976: Small Mammals: Their Productiv'ityDocument1 pageNature Vol. 259 January 29 1976: Small Mammals: Their Productiv'ityKo KaNo ratings yet
- BIOLS340. Chapter 16 SummaryDocument10 pagesBIOLS340. Chapter 16 SummaryNawaf Al.RiffaiNo ratings yet
- Chapter 15 Questions and AnswersDocument3 pagesChapter 15 Questions and AnswersMendes AbeNo ratings yet
- A. O. WEESE.1919.Reacciones Ambientales Del PhrynosomaDocument22 pagesA. O. WEESE.1919.Reacciones Ambientales Del PhrynosomaMarcos Valentin Laurean HernándezNo ratings yet
- Species Abundance and Diversity in the Sonoran DesertDocument2 pagesSpecies Abundance and Diversity in the Sonoran DesertumbuNo ratings yet
- World Distribution, Diversity and Endemism of Aquatic MacrophytesDocument16 pagesWorld Distribution, Diversity and Endemism of Aquatic MacrophytesJonathan Urrutia EstradaNo ratings yet
- The Anatomy of A 'Suture Zone' in Amazonian Butter Ies: A Coalescent-Based Test For Vicariant Geographic Divergence and SpeciationDocument20 pagesThe Anatomy of A 'Suture Zone' in Amazonian Butter Ies: A Coalescent-Based Test For Vicariant Geographic Divergence and SpeciationDaniel RodriguezNo ratings yet
- Impact of Exotic Pastures On Epigeic Arthropod Diversity and Contribution of Native and Exotic PlantDocument10 pagesImpact of Exotic Pastures On Epigeic Arthropod Diversity and Contribution of Native and Exotic PlantDomingas RosaNo ratings yet
- Biome - WikipediaDocument70 pagesBiome - WikipediaPran pynNo ratings yet
- Revista Mexicana de Biodiversidad 78: 303 - 310, 2007Document8 pagesRevista Mexicana de Biodiversidad 78: 303 - 310, 2007ElyRangelNo ratings yet
- Estimating Growth, Phenology and Water Balance of Annual Mediterranean Pastures Using a Simple Simulation ModelDocument13 pagesEstimating Growth, Phenology and Water Balance of Annual Mediterranean Pastures Using a Simple Simulation ModelCristóbal Alberto Campos MuñozNo ratings yet
- Cracraft 1985 Aves-EndemismosDocument37 pagesCracraft 1985 Aves-EndemismosAndrigo Ulian AgostiniNo ratings yet
- Review VarillalesDocument18 pagesReview Varillalesjhoan andrey beltranNo ratings yet
- Human Impact on EnvironmentDocument10 pagesHuman Impact on EnvironmentkandefrancisNo ratings yet
- Exercises Have To Be UsedDocument3 pagesExercises Have To Be UsedRosalinda MintreNo ratings yet
- Noaa 34225 DS1Document15 pagesNoaa 34225 DS1Luis Ángel Martínez GálvezNo ratings yet
- Phycology For The Mammalogist: Marine Rocky Shores and Mammal-Dominated Communities-How Different Are The Structuring Processes?Document12 pagesPhycology For The Mammalogist: Marine Rocky Shores and Mammal-Dominated Communities-How Different Are The Structuring Processes?davidNo ratings yet
- Bouvy 2000 Occurrence of Cylindrospermopsis (Cyanobacteria) in 39 Brazilian Tropical Reservoirs DDocument15 pagesBouvy 2000 Occurrence of Cylindrospermopsis (Cyanobacteria) in 39 Brazilian Tropical Reservoirs DSal TavaresNo ratings yet
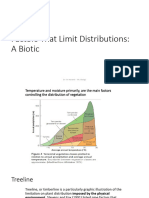
- Week - 5. Factors That Limit Distributions.a Biotic. NicheDocument31 pagesWeek - 5. Factors That Limit Distributions.a Biotic. NicheAnnisa VeterinerianNo ratings yet
- 1 s2.0 S0038071721001541 MainDocument11 pages1 s2.0 S0038071721001541 MainJosé Ribeiro JuniorNo ratings yet
- Ecological succession and ecosystem resilienceDocument4 pagesEcological succession and ecosystem resilienceJuan José Zapata FrancoNo ratings yet
- SHIFT Project Soil Ants Final ReportDocument7 pagesSHIFT Project Soil Ants Final ReportmvbgarciaNo ratings yet
- Zoologia Article 30475 en 1Document9 pagesZoologia Article 30475 en 1Angela Maria Mendivelso TaracheNo ratings yet
- Garcia. 2018. The Biodiversity of Small Passerine Birds at Wetlands - Species Loss and Effects of Climate ChangeDocument5 pagesGarcia. 2018. The Biodiversity of Small Passerine Birds at Wetlands - Species Loss and Effects of Climate ChangeMauricio Chávez ParraNo ratings yet
- Jordan 3e Chapter3 PDFDocument20 pagesJordan 3e Chapter3 PDFShane Franchezka GarciaNo ratings yet
- An Introduction To The Ecology of Desert SystemsDocument28 pagesAn Introduction To The Ecology of Desert SystemsInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Longino Coddington Colwell 2002Document14 pagesLongino Coddington Colwell 2002Aline NeNo ratings yet
- BIS 2B Midterm 1 Fall 2011 Study GuideDocument22 pagesBIS 2B Midterm 1 Fall 2011 Study GuideKinpopNo ratings yet
- Journal of Arid Environments: Leonardo D. Fern AndezDocument5 pagesJournal of Arid Environments: Leonardo D. Fern AndezRj EfogonNo ratings yet
- Missouri Botanical Garden PressDocument34 pagesMissouri Botanical Garden PressAriadne Cristina De AntonioNo ratings yet
- Extractive Logicofthe Colonialityof Nature Feeling Thinking Through Agroecologyasa Decolonial ProjectDocument20 pagesExtractive Logicofthe Colonialityof Nature Feeling Thinking Through Agroecologyasa Decolonial ProjectVictoria PérezNo ratings yet
- Sustainability 14 02881Document13 pagesSustainability 14 02881Victoria PérezNo ratings yet
- Havkin-Frenkel, Daphna Belanger, Faith C. (2018) - Handbook of Vanilla Science and Technology. Vanilla Diseases.Document14 pagesHavkin-Frenkel, Daphna Belanger, Faith C. (2018) - Handbook of Vanilla Science and Technology. Vanilla Diseases.Victoria PérezNo ratings yet
- Vidal, O., & Brusca, R. C. (2020) - Mexico's Biocultural Diversity in Peril.Document23 pagesVidal, O., & Brusca, R. C. (2020) - Mexico's Biocultural Diversity in Peril.Victoria PérezNo ratings yet
- Nhamo, G. (2012) - Participatory Action Research As A Platform For Community Engagement in Higher EducationDocument22 pagesNhamo, G. (2012) - Participatory Action Research As A Platform For Community Engagement in Higher EducationVictoria PérezNo ratings yet
- Environment Perceptions and Conservation PreferencesDocument10 pagesEnvironment Perceptions and Conservation PreferencesVictoria PérezNo ratings yet
- Victor Toledo ArticuloDocument17 pagesVictor Toledo ArticuloLaura TrujilloNo ratings yet
- Perceptions Regarding The Challenges and Constraints Faced by Smallholder Farmers of Vanilla in MexicoDocument24 pagesPerceptions Regarding The Challenges and Constraints Faced by Smallholder Farmers of Vanilla in MexicoVictoria PérezNo ratings yet
- Spirit Bear LessonDocument2 pagesSpirit Bear LessonmichellemillageNo ratings yet
- Basic Techniques in Technical WritingDocument5 pagesBasic Techniques in Technical Writingdave vincin gironNo ratings yet
- What is Biodiversity? Exploring Genetic, Species and Ecosystem DiversityDocument4 pagesWhat is Biodiversity? Exploring Genetic, Species and Ecosystem DiversityBella EstherNo ratings yet
- Chapter X: Air, Water and Soil Resources: Air As A Natural ResourceDocument44 pagesChapter X: Air, Water and Soil Resources: Air As A Natural ResourceAudrey Maree Anoche AdraNo ratings yet
- Paper 16 Vinay Malik PDFDocument5 pagesPaper 16 Vinay Malik PDFPRABHAT KUMAR CHAUHANNo ratings yet
- GE01 AssignmentsDocument3 pagesGE01 AssignmentsFranco HaganNo ratings yet
- W11 Biodiversity GmoDocument23 pagesW11 Biodiversity GmoJerald BacacaoNo ratings yet
- Edsc Weekly Schedule Format w2 FinalDocument3 pagesEdsc Weekly Schedule Format w2 Finalapi-287882801No ratings yet
- Darwin's Influence On Modern ThoughtDocument6 pagesDarwin's Influence On Modern ThoughtsandarshreddyNo ratings yet
- Wegener's Continental Drift Theory challenged scientific establishmentDocument6 pagesWegener's Continental Drift Theory challenged scientific establishmentsiddiqueaquibNo ratings yet
- Tolypothrix DistortaDocument13 pagesTolypothrix DistortaIrinaSalariNo ratings yet
- Why Cross-Cultural Psychology Is Incomplete Without Comparative and Developmental PerspectivesDocument13 pagesWhy Cross-Cultural Psychology Is Incomplete Without Comparative and Developmental PerspectivesbNo ratings yet
- CompartmentalizationDocument4 pagesCompartmentalizationzainab maqboolNo ratings yet
- ZL-101 Biosystematics and TaxonomyDocument99 pagesZL-101 Biosystematics and TaxonomySamvit PatilNo ratings yet
- Online ENVR 1401-Population Tagging Lab Updated 10 - 6 - 2020Document3 pagesOnline ENVR 1401-Population Tagging Lab Updated 10 - 6 - 2020UrielNo ratings yet
- Exercícios: 1. (UNIFOR CE/2006)Document3 pagesExercícios: 1. (UNIFOR CE/2006)erica31No ratings yet
- Class Hours:: Gene 205 Ecology and Evolutionary Biology (2190205)Document3 pagesClass Hours:: Gene 205 Ecology and Evolutionary Biology (2190205)Metehan KaraNo ratings yet
- Bengaluru - Citizenmatters - in - How To Make Mini Forest Miyawaki Method 34867 PDFDocument9 pagesBengaluru - Citizenmatters - in - How To Make Mini Forest Miyawaki Method 34867 PDFDebi GhoshNo ratings yet
- Natural Selection & EvolutionDocument9 pagesNatural Selection & EvolutionDaniel BerryNo ratings yet
- Taxonomy and SystematicsDocument22 pagesTaxonomy and SystematicsHyun JiNo ratings yet
- Foundations of Human EcologyDocument22 pagesFoundations of Human EcologyBo ColemanNo ratings yet
- Science, Technology, and Society Lecture 5: Biodiversity and Human SocietyDocument4 pagesScience, Technology, and Society Lecture 5: Biodiversity and Human Societyjennylyn karunungan100% (1)
- Mca Sir James Mitchell TreesDocument26 pagesMca Sir James Mitchell Treesaimofdesign100% (1)
- Evolution Evidence ExplainedDocument13 pagesEvolution Evidence ExplainedJohn Bernard VeloriaNo ratings yet
- 15 Beaks of FinchesDocument8 pages15 Beaks of FinchesKevin LiNo ratings yet