You might also like
- An Overview About The Biology of Skeletal Muscle Satellite CellsDocument14 pagesAn Overview About The Biology of Skeletal Muscle Satellite CellsCris LiftsNo ratings yet
- The Functions of Kinesin and Kinesin Related Proteins in EukaryotesDocument15 pagesThe Functions of Kinesin and Kinesin Related Proteins in Eukaryotes戴翰明No ratings yet
- Matis, 2019Document7 pagesMatis, 2019MarcoNo ratings yet
- Gardel JCB 2015Document3 pagesGardel JCB 2015useretailNo ratings yet
- 1 PDFDocument20 pages1 PDFOtnellNo ratings yet
- Articulo. La Termodinámica Del Desarrollo en El Diseño de Tejidos BioartificialesDocument11 pagesArticulo. La Termodinámica Del Desarrollo en El Diseño de Tejidos BioartificialesFernanda DíazNo ratings yet
- Senior High School Department Curriculum MapDocument5 pagesSenior High School Department Curriculum MapCamille Sison-AlmirolNo ratings yet
- BP 2Document26 pagesBP 2Kaixian LiuNo ratings yet
- Cytoskeletal Elements in Bacteria: Peter L. GraumannDocument30 pagesCytoskeletal Elements in Bacteria: Peter L. GraumannProcariontesNo ratings yet
- Endless Forms Most Beautiful 2 Teleonomy and The Bioengineering of Chimaeric and Synthetic OrganismsDocument30 pagesEndless Forms Most Beautiful 2 Teleonomy and The Bioengineering of Chimaeric and Synthetic OrganismsYael AracennNo ratings yet
- Topic 3.9 To 3.14: Year: 12 Subject: Biology Unit: 3Document58 pagesTopic 3.9 To 3.14: Year: 12 Subject: Biology Unit: 3reyaNo ratings yet
- Jurnal Integumen 4Document12 pagesJurnal Integumen 4Ummu AthiyyahNo ratings yet
- Bershandsky Et Al. (2006) - Adhesion-Mediated Mechanosensitivity A Time To Experiment, and A Time To TheorizeDocument10 pagesBershandsky Et Al. (2006) - Adhesion-Mediated Mechanosensitivity A Time To Experiment, and A Time To TheorizeTom StevensNo ratings yet
- PHA DENT-Cytoskeleton Cell-Cell ConnectionDocument79 pagesPHA DENT-Cytoskeleton Cell-Cell ConnectionZeynep ZareNo ratings yet
- Cytoskeleton - 2023 - Kerivan - Molecular and Cellular Level Characterization of Cytoskeletal Mechanics Using A QuartzDocument12 pagesCytoskeleton - 2023 - Kerivan - Molecular and Cellular Level Characterization of Cytoskeletal Mechanics Using A Quartzzakariaamehdouk141No ratings yet
- Mitosis: Introduction: Cell CycleDocument3 pagesMitosis: Introduction: Cell CycleIzza Aulia Michellia AlbaNo ratings yet
- 1.6 Cell DivisionDocument46 pages1.6 Cell DivisionJoanita Jasmine JeniferNo ratings yet
- Regulación Biomecánica de La Contractilidad Control Espacial y DinámicaDocument21 pagesRegulación Biomecánica de La Contractilidad Control Espacial y DinámicaKarina B Hernandez ANo ratings yet
- Cellular Nanomechanics 2Document27 pagesCellular Nanomechanics 2Ahmed MotasimNo ratings yet
- Types of Meristematic TissueDocument3 pagesTypes of Meristematic Tissuejagan mohan rao vanaNo ratings yet
- Getting Into Shape: How Do Rod-Like Bacteria Control Their Geometry?Document9 pagesGetting Into Shape: How Do Rod-Like Bacteria Control Their Geometry?FFWW WWNo ratings yet
- LAS - PROTEINS - Franco, Amber L. - 12-CalielDocument2 pagesLAS - PROTEINS - Franco, Amber L. - 12-CalielJl LeeNo ratings yet
- Ow Functional Differentiation Originated in Prebiotic EvolutionDocument23 pagesOw Functional Differentiation Originated in Prebiotic EvolutionVasco De NogalesNo ratings yet
- Adhesion y Migración Celular 2021-1 - CompressedDocument44 pagesAdhesion y Migración Celular 2021-1 - CompressedNiyuhNo ratings yet
- Regulation of The Meiotic Divisions of Mammalian Oocytes and EggsDocument10 pagesRegulation of The Meiotic Divisions of Mammalian Oocytes and EggsAlexandra CardosoNo ratings yet
- MorphogenesisDocument25 pagesMorphogenesisDrAbhilasha SharmaNo ratings yet
- Self-Assembly of Proteins: Dr. K. Uma Maheswari Professor, School of Chemical & Biotechnology SASTRA UniversityDocument10 pagesSelf-Assembly of Proteins: Dr. K. Uma Maheswari Professor, School of Chemical & Biotechnology SASTRA University18UGBT055 Surya PrabhaNo ratings yet
- Cell Organelles - Structure and Functions of Cell OrganellesDocument18 pagesCell Organelles - Structure and Functions of Cell OrganellesUnza WaqarNo ratings yet
- Structural Organization of Life Chapter SummaryDocument74 pagesStructural Organization of Life Chapter SummaryParkash Kumar RathoreNo ratings yet
- Self-Organization in Cellular ArchitectureDocument5 pagesSelf-Organization in Cellular ArchitectureRazvan ZamfiraNo ratings yet
- Cell, Basic Structure of All Living Creatures: Life Science, Human BiologyDocument11 pagesCell, Basic Structure of All Living Creatures: Life Science, Human BiologyAshleigh Jenson WilliamsonNo ratings yet
- Advances in 3D multicellular spheroids formationDocument15 pagesAdvances in 3D multicellular spheroids formationDaniela Mădălina GhețuNo ratings yet
- Reviews: Mechanomicrobiology: How Bacteria Sense and Respond To ForcesDocument14 pagesReviews: Mechanomicrobiology: How Bacteria Sense and Respond To ForcesOded SabahNo ratings yet
- HHS Public Access: Mitochondrial Fission and FusionDocument15 pagesHHS Public Access: Mitochondrial Fission and FusionPramod KcNo ratings yet
- Life - PaperDocument32 pagesLife - PaperPrashant VemuriNo ratings yet
- BioEssays - 2002 - Tyson - The Dynamics of Cell Cycle RegulationDocument15 pagesBioEssays - 2002 - Tyson - The Dynamics of Cell Cycle RegulationNindya AgustiaNo ratings yet
- Mitosis MeiosisDocument16 pagesMitosis MeiosisImmanuel MarcusNo ratings yet
- Activity No. 3: Structure of A Cell and MitosisDocument9 pagesActivity No. 3: Structure of A Cell and MitosisEricka GenoveNo ratings yet
- xenobots2Document14 pagesxenobots2yaswanthjetti21No ratings yet
- Bies 201900157Document9 pagesBies 201900157Paulius SungailaNo ratings yet
- Bruckner 2015. AFM Over EpitheliaDocument8 pagesBruckner 2015. AFM Over EpitheliaCristina Pinto DueñasNo ratings yet
- Act. 3 Cytology PeraltaDocument18 pagesAct. 3 Cytology PeraltaCogie PeraltaNo ratings yet
- Tissue Engineering: Name: Frankie Jonathan Laloo ROLL NO:1378 M.Sc. (Final)Document7 pagesTissue Engineering: Name: Frankie Jonathan Laloo ROLL NO:1378 M.Sc. (Final)Anurag PanchalNo ratings yet
- Mitochondrion Related Annurev - Micro.62.081307Document24 pagesMitochondrion Related Annurev - Micro.62.081307Paulius SungailaNo ratings yet
- SHS STEM Bio1 Q1 Week 4 Module 8 - Significance or Applications of Mitosis and MeiosisDocument17 pagesSHS STEM Bio1 Q1 Week 4 Module 8 - Significance or Applications of Mitosis and MeiosisEmer Perez100% (1)
- Perspectives: A Road Map For The Development of Community Systems (Cosy) BiologyDocument7 pagesPerspectives: A Road Map For The Development of Community Systems (Cosy) BiologyandreaNo ratings yet
- 1.6 Cell DivisionDocument30 pages1.6 Cell DivisionAnnabeth ParkerNo ratings yet
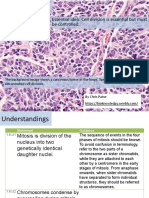
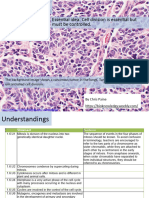
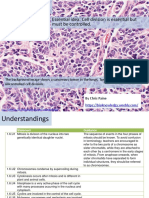
- 1.6 Cell Division: Essential Idea: Cell Division Is Essential But Must Be ControlledDocument33 pages1.6 Cell Division: Essential Idea: Cell Division Is Essential But Must Be ControlledAnonymous bOMSH4wDgeNo ratings yet
- Biophysical stimuli influence cell differentiation during tissue formationDocument10 pagesBiophysical stimuli influence cell differentiation during tissue formationBastion ReinhartNo ratings yet
- Genetics Lesson 1Document12 pagesGenetics Lesson 1VK VillNo ratings yet
- Cell Cycle - Mitosis and Meiosis PDFDocument43 pagesCell Cycle - Mitosis and Meiosis PDFVEENA DEVINo ratings yet
- Cells: Building Blocks of LifeDocument76 pagesCells: Building Blocks of LifeMilka RahmanNo ratings yet
- Artificial Cells From Basic Science To ApplicationDocument17 pagesArtificial Cells From Basic Science To ApplicationNo Speed LimitNo ratings yet
- GENERAL BIOLOGY 1: THE STUDY OF LIFEDocument25 pagesGENERAL BIOLOGY 1: THE STUDY OF LIFEJuanito MerciNo ratings yet
- 2020, "Looping In" Mechanics - Mechanobiologic Regulation of The Nucleus and The EpigenomeDocument26 pages2020, "Looping In" Mechanics - Mechanobiologic Regulation of The Nucleus and The EpigenomeErnesto RojoNo ratings yet
- EpidemiologyDocument21 pagesEpidemiologyAgustina TutiNo ratings yet
- Activity 4 - The Cell CycleDocument9 pagesActivity 4 - The Cell CyclejaNo ratings yet
- The Extracellular Matrix in Development PDFDocument16 pagesThe Extracellular Matrix in Development PDFMaura HokamaNo ratings yet
- The Actin Cortex at A GlanceDocument9 pagesThe Actin Cortex at A GlanceArkayan LahaNo ratings yet
- Gerkin Et Al 2018 Towards Systematic Data Driven Validation of A Collaborative Multi Scale Model of CaenorhabditisDocument8 pagesGerkin Et Al 2018 Towards Systematic Data Driven Validation of A Collaborative Multi Scale Model of CaenorhabditisVasillis MamosNo ratings yet
- What Makes The Maternal X Chromosome Resistant To Undergoing Imprinted X Inactivation?Document5 pagesWhat Makes The Maternal X Chromosome Resistant To Undergoing Imprinted X Inactivation?Vasillis MamosNo ratings yet
- Impacts of Extreme Summers On European Ecosystems: A Comparative Analysis of 2003, 2010 and 2018Document10 pagesImpacts of Extreme Summers On European Ecosystems: A Comparative Analysis of 2003, 2010 and 2018Vasillis MamosNo ratings yet
- Evolution of Mechanisms and Behaviour Important For PainDocument8 pagesEvolution of Mechanisms and Behaviour Important For PainVasillis MamosNo ratings yet
- Polycomb Complexes in X Chromosome InactivationDocument5 pagesPolycomb Complexes in X Chromosome InactivationVasillis MamosNo ratings yet
- New Ancient Eastern European Yersinia Pestis Genomes Illuminate The Dispersal of Plague in EuropeDocument8 pagesNew Ancient Eastern European Yersinia Pestis Genomes Illuminate The Dispersal of Plague in EuropeVasillis MamosNo ratings yet
- Altered Energy Partitioning Across Terrestrial Ecosystems in The European Drought Year 2018Document8 pagesAltered Energy Partitioning Across Terrestrial Ecosystems in The European Drought Year 2018Vasillis MamosNo ratings yet
- Pain in Dinosaurs: What Is The Evidence?Document8 pagesPain in Dinosaurs: What Is The Evidence?Vasillis MamosNo ratings yet
- An Ecometric Analysis of The Fossil Mammal Record of The Turkana BasinDocument13 pagesAn Ecometric Analysis of The Fossil Mammal Record of The Turkana BasinVasillis MamosNo ratings yet
- Are Some Cultures More Mind-Minded in Their Moral Judgements Than Others?Document8 pagesAre Some Cultures More Mind-Minded in Their Moral Judgements Than Others?Vasillis MamosNo ratings yet
- The Origin and Evolution of Homo SapiensDocument12 pagesThe Origin and Evolution of Homo SapiensThiago S. Sales0% (1)
- Coordination Between Patterning and Morphogenesis Ensures Robustness During Mouse DevelopmentDocument12 pagesCoordination Between Patterning and Morphogenesis Ensures Robustness During Mouse DevelopmentVasillis MamosNo ratings yet
- Functional Diversity of Microbial Ecologies Estimated From Ancient Human Coprolites and Dental CalculusDocument13 pagesFunctional Diversity of Microbial Ecologies Estimated From Ancient Human Coprolites and Dental CalculusVasillis MamosNo ratings yet
- New Ancient Eastern European Yersinia Pestis Genomes Illuminate The Dispersal of Plague in EuropeDocument8 pagesNew Ancient Eastern European Yersinia Pestis Genomes Illuminate The Dispersal of Plague in EuropeVasillis MamosNo ratings yet
- Pergamon: Z Beha ', Ther. & Exp. PsychiatDocument4 pagesPergamon: Z Beha ', Ther. & Exp. PsychiatVasillis MamosNo ratings yet
- Does Epigenetic Memory' of Early-Life Stress Predispose To Chronic Pain in Later Life? A Potential Role For The Stress Regulator FKBP5Document8 pagesDoes Epigenetic Memory' of Early-Life Stress Predispose To Chronic Pain in Later Life? A Potential Role For The Stress Regulator FKBP5Vasillis MamosNo ratings yet
- Pergamon: The Sensory OrderDocument4 pagesPergamon: The Sensory OrderVasillis MamosNo ratings yet
- Pergamon: The) The)Document8 pagesPergamon: The) The)Vasillis MamosNo ratings yet
- Self-.Efficacy: A Predictor But Not A Cause of BehaviorDocument6 pagesSelf-.Efficacy: A Predictor But Not A Cause of BehaviorVasillis MamosNo ratings yet
- EysenckDocument11 pagesEysenckLuisNo ratings yet
- Hawkins 1995Document6 pagesHawkins 1995Vasillis MamosNo ratings yet
- Self-.Efficacy: A Predictor But Not A Cause of BehaviorDocument6 pagesSelf-.Efficacy: A Predictor But Not A Cause of BehaviorVasillis MamosNo ratings yet
- The New Brain Sciences Perils and Prospects by Dai Rees, Steven RoseDocument317 pagesThe New Brain Sciences Perils and Prospects by Dai Rees, Steven RoseVasillis MamosNo ratings yet
- On Curvature and Torsion in Courant Algebroids, Paolo Aschieri, Francesco Bonechi, Andreas DeserDocument27 pagesOn Curvature and Torsion in Courant Algebroids, Paolo Aschieri, Francesco Bonechi, Andreas DeserVasillis MamosNo ratings yet
- Transdisciplinarity TJESNo 1 12 2010Document20 pagesTransdisciplinarity TJESNo 1 12 2010비단터No ratings yet
- Symmetry of Superconducting Correlations in Displaced Bilayers of Graphene, Mohammad Alidoust, Morten Willatzen and Antti-Pekka JauhoDocument16 pagesSymmetry of Superconducting Correlations in Displaced Bilayers of Graphene, Mohammad Alidoust, Morten Willatzen and Antti-Pekka JauhoVasillis MamosNo ratings yet
- Bandura 1995Document12 pagesBandura 1995Vasillis MamosNo ratings yet
- Dr. Andreas Ipp, Simulations of The Glasma in 3+1DDocument189 pagesDr. Andreas Ipp, Simulations of The Glasma in 3+1DVasillis MamosNo ratings yet
- Will M Gervais - The Zeus Problem Why Representational Content Biases Cannot Explain Faith in GodsDocument8 pagesWill M Gervais - The Zeus Problem Why Representational Content Biases Cannot Explain Faith in GodsAdam RauhNo ratings yet
- Bone and CartilageDocument56 pagesBone and CartilageGodwin ZengeniNo ratings yet
- Heart Health GuideDocument7 pagesHeart Health GuideYAO CESAR KOFFINo ratings yet
- Science 4 Lesson PlanDocument11 pagesScience 4 Lesson PlanEmily BucsitNo ratings yet
- Muscular System MindmapDocument1 pageMuscular System MindmapMayeliz UrvinaNo ratings yet
- Animal Tissues - DPP 2.1 - (CDS - 2 Viraat 2023)Document3 pagesAnimal Tissues - DPP 2.1 - (CDS - 2 Viraat 2023)tanwar9780No ratings yet
- M 1 PPT-PDF XI Bio CH 7 Structural Organisation in AnimalsDocument39 pagesM 1 PPT-PDF XI Bio CH 7 Structural Organisation in Animalsamarendrar87No ratings yet
- GIT2Document18 pagesGIT2Yasin AbassNo ratings yet
- Cytology, Histology, EmbryologyDocument53 pagesCytology, Histology, Embryologyagostino.93.sommaNo ratings yet
- Module 5 - Body TissuesDocument10 pagesModule 5 - Body Tissues10. Briol AlvinNo ratings yet
- Lipoma of The Oral and Maxillofacial RegionDocument10 pagesLipoma of The Oral and Maxillofacial RegionLaila FortunatoNo ratings yet
- PPT-6 Transportation and Excretion in PlantsDocument19 pagesPPT-6 Transportation and Excretion in PlantskochoaarakochoNo ratings yet
- Anaphy Chap 1-12Document103 pagesAnaphy Chap 1-12Margaret Isabel LacdaoNo ratings yet
- Roni - Support and Locomotion SolutionsDocument13 pagesRoni - Support and Locomotion SolutionsDaniel Mark OgwalNo ratings yet
- Models: Models of CellsDocument10 pagesModels: Models of CellsSolomon SohNo ratings yet
- 1 Respiratory SystemDocument70 pages1 Respiratory SystemJan IceNo ratings yet
- Lesson 1 Comparative AnatomyDocument6 pagesLesson 1 Comparative AnatomyYu Lin100% (1)
- Essentials of Medical PhysiologyDocument6 pagesEssentials of Medical Physiologykumarraushan1000No ratings yet
- Buccal Smear Lab ReportDocument6 pagesBuccal Smear Lab ReportAugustNo ratings yet
- Head and Neck Part II Spotters QuestionsDocument26 pagesHead and Neck Part II Spotters Questions76 S.SHRAVAN KUMARNo ratings yet
- Early Development and Axis Formation in Amphibians and FishDocument3 pagesEarly Development and Axis Formation in Amphibians and FishJohnkevin NocheNo ratings yet
- Dactyloscopy 1st Prelim Exam.Document3 pagesDactyloscopy 1st Prelim Exam.Ocyub Avlas OdnamraNo ratings yet
- The Digestive and Excretory SystemDocument16 pagesThe Digestive and Excretory SystemMiguel Angel Loor MolinaNo ratings yet
- Introduction To Anatomy and Physiology: Unit IDocument55 pagesIntroduction To Anatomy and Physiology: Unit ILara C.No ratings yet
- Periodontal LigamentDocument27 pagesPeriodontal Ligamentsamar yousif mohamedNo ratings yet
- Iam Free Jan'24 MRCS ADocument4 pagesIam Free Jan'24 MRCS AthanitoNo ratings yet
- Introduction To The Body As A WholeDocument29 pagesIntroduction To The Body As A Wholekhizer hayatNo ratings yet
- NCERT Bio FactsDocument7 pagesNCERT Bio FactsGomathi MgmNo ratings yet
- Assessment Test in Science 3.1Document2 pagesAssessment Test in Science 3.1Caren Mae PunzalanNo ratings yet
- Chapter 7 - Muscular SystemDocument50 pagesChapter 7 - Muscular SystemZea Mae AgirNo ratings yet
- Standardized Identification Points For Histology Slides ForDocument29 pagesStandardized Identification Points For Histology Slides ForAroosha JamshaidNo ratings yet