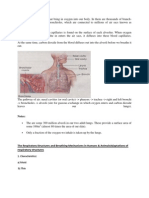
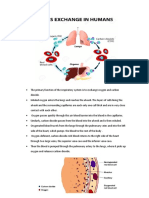
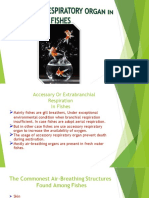
You might also like
- Osteichthyes PDFDocument50 pagesOsteichthyes PDFmuhammadismailNo ratings yet
- Answer Keys Life 2 WBDocument18 pagesAnswer Keys Life 2 WBClara PenínNo ratings yet
- Scope 020114 Nonfiction SharksDocument5 pagesScope 020114 Nonfiction Sharksapi-206219067No ratings yet
- Mermaid Magic Connecting With The Energy of The Ocean and The Healing Power of Water (Serene Conneeley Lucy Cavendish) (Z-Library)Document417 pagesMermaid Magic Connecting With The Energy of The Ocean and The Healing Power of Water (Serene Conneeley Lucy Cavendish) (Z-Library)MysticNo ratings yet
- The Maritime Foundations of Andean CivilizationDocument10 pagesThe Maritime Foundations of Andean CivilizationMiguel Castillo RodriguezNo ratings yet
- National Geographic UK - June 2023Document130 pagesNational Geographic UK - June 2023ALESSANDRO PREMOLINo ratings yet
- Respiratory AssessmentDocument12 pagesRespiratory AssessmentAmanda ScarletNo ratings yet
- Evolution of Respiratory SystemDocument10 pagesEvolution of Respiratory SystemWwwanand111No ratings yet
- Lightening Organs of FishDocument22 pagesLightening Organs of FishHeer SheikhNo ratings yet
- Air Breathing OrgansDocument13 pagesAir Breathing OrgansS. JeroNo ratings yet
- Our Lungs Are The Organs That Bring in Oxygen Into Our BodyDocument17 pagesOur Lungs Are The Organs That Bring in Oxygen Into Our BodySaidatul Atyah Mohd ApendaiNo ratings yet
- Respiratory System: Lecture Six Assit - Lect.Maha MustafaDocument8 pagesRespiratory System: Lecture Six Assit - Lect.Maha Mustafanareman hassanNo ratings yet
- Gas Exchange Written ReportDocument11 pagesGas Exchange Written ReportEllanny GloriaNo ratings yet
- Accessory Respiratory Organ in FishDocument47 pagesAccessory Respiratory Organ in Fishkoustav kundu0% (1)
- ApbioprojectDocument5 pagesApbioprojectAshleyNo ratings yet
- Respiratory System Insects 2Document4 pagesRespiratory System Insects 2KIPNGETICH FREDRICKNo ratings yet
- Reptilian Lungs: Reptile Anatomy#respirationDocument1 pageReptilian Lungs: Reptile Anatomy#respirationa2962109No ratings yet
- Chapter 9 Respiratory Systems NotesDocument11 pagesChapter 9 Respiratory Systems NotesCaleb DodgeNo ratings yet
- Comparative Anotomy of Respiratory SystemDocument12 pagesComparative Anotomy of Respiratory SystemNarasimha MurthyNo ratings yet
- Breathing and Exchange of GasesDocument17 pagesBreathing and Exchange of GasesArjun ChristopherNo ratings yet
- Respiratory System PreDocument11 pagesRespiratory System PrecontehamosaNo ratings yet
- 156 P18ZC101 2020120606191995Document15 pages156 P18ZC101 2020120606191995prachiteegoregaonkarNo ratings yet
- 4th Q Module GEN BIO 2 Week 5 6Document19 pages4th Q Module GEN BIO 2 Week 5 6Hannah CastroNo ratings yet
- Chapter 8Document14 pagesChapter 8Shatviga VisvalingamNo ratings yet
- Respiratory System-Chapter 17Document7 pagesRespiratory System-Chapter 17Yza S.No ratings yet
- CirculatoryDocument12 pagesCirculatoryjhosepNo ratings yet
- Respiratory System: Respiration (Oxidation of Food in Cells Cellular Respiration Energy Is Produced Along With WaterDocument15 pagesRespiratory System: Respiration (Oxidation of Food in Cells Cellular Respiration Energy Is Produced Along With WaterShuvanjan DahalNo ratings yet
- Anatomy and Physiology of The Respiratory System (Vizconde, Ehreiz Raiden C. BSN2A)Document4 pagesAnatomy and Physiology of The Respiratory System (Vizconde, Ehreiz Raiden C. BSN2A)Raiden VizcondeNo ratings yet
- Theory of LungDocument7 pagesTheory of LungAndres Dario R LozNo ratings yet
- Gas Exchange in AnimalDocument3 pagesGas Exchange in AnimalAaron Dave CanteroNo ratings yet
- Gas Exchange in AnimalDocument3 pagesGas Exchange in AnimalAaron Dave CanteroNo ratings yet
- Breathing and ExchangeDocument40 pagesBreathing and ExchangeShambhaviNo ratings yet
- Resprn in Humns-KDBDocument52 pagesResprn in Humns-KDBKiranNo ratings yet
- The Respiratory SystemDocument8 pagesThe Respiratory SystemDavid DanielNo ratings yet
- Respiratory Structures in Pila:: Aquatic RespirationDocument8 pagesRespiratory Structures in Pila:: Aquatic Respirationazizi farhanNo ratings yet
- 2.D.2 Homeostatic MechanismsDocument42 pages2.D.2 Homeostatic Mechanismsapi-373344212No ratings yet
- Respiratory SystemDocument3 pagesRespiratory Systemๆรำำทแร หบหฝำมฟNo ratings yet
- Notes 1Document12 pagesNotes 1Muhammad TaimoorNo ratings yet
- Anatomy and Physiology of PCAPDocument8 pagesAnatomy and Physiology of PCAPMads0% (1)
- Human Anatomy Reading ComprehensionDocument16 pagesHuman Anatomy Reading ComprehensionRhams BairullaNo ratings yet
- For Other Uses, See - "Lunged" Redirects Here. For Other Uses, SeeDocument12 pagesFor Other Uses, See - "Lunged" Redirects Here. For Other Uses, Seefifa92_piscesNo ratings yet
- Class 7 RespirationDocument5 pagesClass 7 RespirationGopi SNo ratings yet
- Handout Respiratory System 2022 2023Document3 pagesHandout Respiratory System 2022 2023reece sayamNo ratings yet
- Anatomy and PhysiologyDocument11 pagesAnatomy and Physiologyjawn09euclidNo ratings yet
- Respiratory System s2 2019Document56 pagesRespiratory System s2 2019noviantyramadhani12No ratings yet
- Ma Nurfaizah Ma04200026 Respiratory System April 2020Document3 pagesMa Nurfaizah Ma04200026 Respiratory System April 2020Nur Faizah Binti Jamail MANo ratings yet
- Respiratory Organ of FrogDocument1 pageRespiratory Organ of Frogkunalkumaryadav086No ratings yet
- PDBreathing and Exchange of Gases Class 11 Notes CBSE Biology Chapter 17FDocument15 pagesPDBreathing and Exchange of Gases Class 11 Notes CBSE Biology Chapter 17Fgandhitirth147No ratings yet
- Breathing Process PDFDocument3 pagesBreathing Process PDFDianeNo ratings yet
- 13 Respiratory SystemDocument5 pages13 Respiratory Systemvada_soNo ratings yet
- OCR (A) Biology A-Level: Topic 3.1: Exchange and TransportDocument4 pagesOCR (A) Biology A-Level: Topic 3.1: Exchange and TransportNguyen Thi Thien MinhNo ratings yet
- Bilology XII 2014 CH - 14 RespirationDocument16 pagesBilology XII 2014 CH - 14 RespirationMuhammad UsmanNo ratings yet
- Gas Exchange in Animals3Document47 pagesGas Exchange in Animals3Ashley JibrielleNo ratings yet
- Accessory Air Breathing OrgansDocument26 pagesAccessory Air Breathing Organskaushik mallickNo ratings yet
- Respiratory SystemDocument9 pagesRespiratory SystemShivani Sriram100% (1)
- Lect 3Document8 pagesLect 3علي الاسديNo ratings yet
- Respiratory PhysiologyDocument15 pagesRespiratory PhysiologySahil SalluNo ratings yet
- Anatomic and Physiologic OverviewDocument8 pagesAnatomic and Physiologic OverviewJoseph King MacaranasNo ratings yet
- Gaseous Exchange A LevelDocument33 pagesGaseous Exchange A LevelemmanuelkeetoNo ratings yet
- BIO 342 Comparative Vertebrate Anatomy Lecture Notes - Respiratory System Respiration Is The Process of Obtaining Oxygen From The External Environment &Document13 pagesBIO 342 Comparative Vertebrate Anatomy Lecture Notes - Respiratory System Respiration Is The Process of Obtaining Oxygen From The External Environment &PearNo ratings yet
- Anatomy of The Respiratory SystemDocument37 pagesAnatomy of The Respiratory SystemPal SedNo ratings yet
- Respiratory SupplementDocument3 pagesRespiratory SupplementSebastian SmytheNo ratings yet
- Respiratory Diseases of Parrots - Anatomy, Physiology, Diagnosis and Treatment - Crosta, L. (2021)Document22 pagesRespiratory Diseases of Parrots - Anatomy, Physiology, Diagnosis and Treatment - Crosta, L. (2021)Veronica Hdez FloresNo ratings yet
- Our Common Insects A Popular Account of the Insects of Our Fields, Forests, Gardens and HousesFrom EverandOur Common Insects A Popular Account of the Insects of Our Fields, Forests, Gardens and HousesNo ratings yet
- The British Woodlice: Being a Monograph of the Terrestrial Isopod Crustacea Occurring in the British IslandsFrom EverandThe British Woodlice: Being a Monograph of the Terrestrial Isopod Crustacea Occurring in the British IslandsNo ratings yet
- Accessory Respiratory Organ-FishDocument63 pagesAccessory Respiratory Organ-Fishkaushik mallickNo ratings yet
- Accessory Respiratory Organ - FishDocument27 pagesAccessory Respiratory Organ - Fishkaushik mallick100% (1)
- Advanced Features of Vertebrates Over ProtochordatesDocument4 pagesAdvanced Features of Vertebrates Over Protochordateskaushik mallickNo ratings yet
- Accessory Air Breathing OrgansDocument26 pagesAccessory Air Breathing Organskaushik mallickNo ratings yet
- Fish Escabeche With Orange (Steamed)Document2 pagesFish Escabeche With Orange (Steamed)itchieNo ratings yet
- Rjecnik HRV EngDocument981 pagesRjecnik HRV EngStevanka ČakmakNo ratings yet
- Fisheries Laws, Policies, and Institutions Answer The Following QuestionsDocument2 pagesFisheries Laws, Policies, and Institutions Answer The Following QuestionsRhea CrisoloNo ratings yet
- Coastal Management Orientation and OverviewDocument25 pagesCoastal Management Orientation and OverviewThessa Vee Capuno RioNo ratings yet
- The Economic Theory of A Common-Property Resource The FisheryDocument6 pagesThe Economic Theory of A Common-Property Resource The FisheryAlbert UgarteNo ratings yet
- Assignments of Fish HabitatDocument2 pagesAssignments of Fish HabitatUmiysNo ratings yet
- Animal Crossing WW CheatsDocument9 pagesAnimal Crossing WW CheatsImogen DoddsNo ratings yet
- English For Fisheries and Marine Science - Unit 2 MUHAMMAD NANDA FIRDAUS - 205070307111019Document7 pagesEnglish For Fisheries and Marine Science - Unit 2 MUHAMMAD NANDA FIRDAUS - 205070307111019Nanda FirdaNo ratings yet
- Mohamed Abdullahi Mohamed "Germany": Promoted byDocument21 pagesMohamed Abdullahi Mohamed "Germany": Promoted byMAslah Kader BoonNo ratings yet
- Takabonerate National ParkDocument15 pagesTakabonerate National ParkIvana Thynaba NarezaNo ratings yet
- Fao Species Identification Sheets Fishing Area 51 (W. Indian Ocean)Document11 pagesFao Species Identification Sheets Fishing Area 51 (W. Indian Ocean)လက္ ေဆာင္No ratings yet
- Acrs2015 Paper-Id 534Document9 pagesAcrs2015 Paper-Id 534Francis Ben Degenion TaglucopNo ratings yet
- Dwnload Full Public Health Nursing Population Centered Health Care in The Community 9th Edition Stanhope Test Bank PDFDocument36 pagesDwnload Full Public Health Nursing Population Centered Health Care in The Community 9th Edition Stanhope Test Bank PDFcosmicalfashionz10zbt89% (9)
- Tilt Cove Newfoundland - in Memory of An Old Mining TownDocument6 pagesTilt Cove Newfoundland - in Memory of An Old Mining Towndb7No ratings yet
- An Analysis of Characterization of Santiago in The Old Man and The SeaDocument10 pagesAn Analysis of Characterization of Santiago in The Old Man and The Seayana latifa100% (1)
- Aral Sea Case StudyDocument18 pagesAral Sea Case StudyMihrican SatışNo ratings yet
- R. M. Ballantyne: The Coral IslandDocument4 pagesR. M. Ballantyne: The Coral IslandValentina VinogradovaNo ratings yet
- Bof 3 Leveling Up GuideDocument61 pagesBof 3 Leveling Up GuideYoas Adhitya RahardjoNo ratings yet
- Lake VictoriaDocument19 pagesLake VictoriaYohans EjiguNo ratings yet
- The Poor Man Who Became King: A Christmas StoryDocument5 pagesThe Poor Man Who Became King: A Christmas StoryRessielyn Kates CaperinaNo ratings yet
- Informational Leaflet No.35 Inland AquacultureDocument41 pagesInformational Leaflet No.35 Inland AquacultureJunior QuinteroNo ratings yet
- Actividad 12 (Corregida)Document7 pagesActividad 12 (Corregida)maricat 126No ratings yet
- Actividad #1 Practica-1Document3 pagesActividad #1 Practica-1DAIRYNo ratings yet
- Laker 13 KayakDocument48 pagesLaker 13 KayakKeith WilliamsNo ratings yet