You might also like
- PNAS 2006 Fagot 17564 7Document4 pagesPNAS 2006 Fagot 17564 7asterix2013No ratings yet
- Skill Mastery Inhibits Adoption of Observed Alternative Solutions Among Chimpanzees (Pan Troglodytes)Document8 pagesSkill Mastery Inhibits Adoption of Observed Alternative Solutions Among Chimpanzees (Pan Troglodytes)Alex DanNo ratings yet
- 2005-Effect of Different IlluminationDocument6 pages2005-Effect of Different IlluminationberilNo ratings yet
- PropoDocument12 pagesPropoapi-295869438No ratings yet
- Miletto Petrazzini Et Al 2019, SymmetryDocument15 pagesMiletto Petrazzini Et Al 2019, SymmetryChristian AgrilloNo ratings yet
- Subjective Shortening: A Model of Pigeons' Memory For Event DurationDocument17 pagesSubjective Shortening: A Model of Pigeons' Memory For Event DurationJacob KantorNo ratings yet
- 2019 Timetable BMS5006Document9 pages2019 Timetable BMS5006amrilmaenbNo ratings yet
- Investigating Habituation To A StimulusDocument10 pagesInvestigating Habituation To A Stimulussandali636100% (5)
- 2.3 Ecology and SamplingDocument28 pages2.3 Ecology and SamplingPhoo Pyae Pyae KyawNo ratings yet
- ZF and CAVEFISHDocument6 pagesZF and CAVEFISHsultanmsajidmNo ratings yet
- Research Paper About SnailsDocument6 pagesResearch Paper About Snailsyhclzxwgf100% (1)
- This Is The Original Experiment Run by BLG144 Students in 2017. Foraging Behaviour of Fruit Flies (Drosophila Melanogaster)Document8 pagesThis Is The Original Experiment Run by BLG144 Students in 2017. Foraging Behaviour of Fruit Flies (Drosophila Melanogaster)Dineth GunasekeraNo ratings yet
- Articulo CristianDocument5 pagesArticulo CristianDaBid Lopez RodriguezNo ratings yet
- Large Number Discrimination in Newborn Fish: Laura Piffer, Maria Elena Miletto Petrazzini, Christian AgrilloDocument6 pagesLarge Number Discrimination in Newborn Fish: Laura Piffer, Maria Elena Miletto Petrazzini, Christian AgrilloChristian AgrilloNo ratings yet
- 2014 - Effects of The Number of Subjects On The Dark Light Preference of ZebrafishDocument7 pages2014 - Effects of The Number of Subjects On The Dark Light Preference of ZebrafishBruno R SantosNo ratings yet
- Experimental Study of Torsion Field Effect On LifetimeDocument2 pagesExperimental Study of Torsion Field Effect On Lifetimeapi-186101394No ratings yet
- Cognition in The WildDocument18 pagesCognition in The WildOriolNo ratings yet
- Novel Proteinaceous Infectious Particles Cause ScrapieDocument10 pagesNovel Proteinaceous Infectious Particles Cause ScrapieMikey HaveyNo ratings yet
- Marine Thesis PDFDocument6 pagesMarine Thesis PDFRaquel Pellicier100% (1)
- (Literature?) : Introduction Must HaveDocument19 pages(Literature?) : Introduction Must Haveapi-295869449No ratings yet
- More Experiences Bigger BrainDocument5 pagesMore Experiences Bigger BrainchristinavlahouNo ratings yet
- Statistical Analysis of Mammal Studies Based On Mammal Sleep DataDocument21 pagesStatistical Analysis of Mammal Studies Based On Mammal Sleep DataMuhammad UsmanNo ratings yet
- Lab Anim 2001 Reeb Whitaker 58 73Document17 pagesLab Anim 2001 Reeb Whitaker 58 73maikaNo ratings yet
- Dissertation ZoologyDocument5 pagesDissertation ZoologyPaperWriterOnlineUK100% (1)
- Age-Related Differences For Duration Discrimination in RatsDocument4 pagesAge-Related Differences For Duration Discrimination in RatsjsaccuzzoNo ratings yet
- Musa Lawan WidaDocument6 pagesMusa Lawan WidaSouleyman AhmadNo ratings yet
- 4 EdgarDocument17 pages4 Edgarmadhura480No ratings yet
- Sliding window correlation reveals behavioral subgroups in aged ratsDocument9 pagesSliding window correlation reveals behavioral subgroups in aged ratsAndrés LiconaNo ratings yet
- Behavioural ProcessesDocument11 pagesBehavioural ProcessesharisankarhsNo ratings yet
- Reading Practice Test 3: Can animals countDocument17 pagesReading Practice Test 3: Can animals countalkan gündoğduNo ratings yet
- Show Me Your Fucking Girlbutt (Asshole)Document9 pagesShow Me Your Fucking Girlbutt (Asshole)Restroom ChemistryNo ratings yet
- Zeb 2012 0758Document16 pagesZeb 2012 0758SATRIO PINANDITONo ratings yet
- IELTS Session 5Document4 pagesIELTS Session 5Sophia PirogNo ratings yet
- A Test of The Many-Eyes Hypothesis (Cockatoo)Document7 pagesA Test of The Many-Eyes Hypothesis (Cockatoo)Sara MeloNo ratings yet
- Zebrafish ArtigoDocument10 pagesZebrafish ArtigoRhael PereiraNo ratings yet
- BehavioralDocument4 pagesBehavioralxingyu92No ratings yet
- Genetics of Behavioral DifferencesDocument12 pagesGenetics of Behavioral DifferencesAditya GalihNo ratings yet
- Chapter 16 - Evaluation of Embryotoxicity Using The Zebrafish ModelDocument9 pagesChapter 16 - Evaluation of Embryotoxicity Using The Zebrafish ModelLuis Humberto Armas RendónNo ratings yet
- Chew 2003Document3 pagesChew 2003I ENo ratings yet
- The History of The Lab Rat Is Full of Scientific Triumphs and Ethical QuandariesDocument7 pagesThe History of The Lab Rat Is Full of Scientific Triumphs and Ethical Quandariesgabriel.nathan.kaufmanNo ratings yet
- Inventions: A Versatile Setup For Measuring Multiple Behavior Endpoints in ZebrafishDocument16 pagesInventions: A Versatile Setup For Measuring Multiple Behavior Endpoints in ZebrafishKarthik ManiNo ratings yet
- Hall 1954Document2 pagesHall 1954yom.moguelNo ratings yet
- Assesment of Mouse Anxiety 2014 Natalia KulesskayaDocument9 pagesAssesment of Mouse Anxiety 2014 Natalia Kulesskayagoutham278No ratings yet
- Ultrasonographic Anatomy of Reproductive Female LeDocument13 pagesUltrasonographic Anatomy of Reproductive Female LeLaily IlmiNo ratings yet
- Object Recognition Test in Mice: Nature Protocol December 2013Document8 pagesObject Recognition Test in Mice: Nature Protocol December 2013erikaNo ratings yet
- Habitambilidad en Calles de BogotaDocument7 pagesHabitambilidad en Calles de BogotaOscar CastañedaNo ratings yet
- Paper 2C ESS Lizard-Evolution G10-2Document17 pagesPaper 2C ESS Lizard-Evolution G10-2SAHARE CANO BADWAM-AlumnoNo ratings yet
- Can Snails Become Habituated To A StimulusDocument2 pagesCan Snails Become Habituated To A StimulusSachitra Wijethunga100% (1)
- Tomotani 2012Document8 pagesTomotani 2012Sebastian PereiraNo ratings yet
- The Environmental Predation Risk (Tree Sparrow)Document6 pagesThe Environmental Predation Risk (Tree Sparrow)Sophany PhaukNo ratings yet
- Measuring anxiety levels in mice using elevated plus mazeDocument3 pagesMeasuring anxiety levels in mice using elevated plus mazeKatherine Herza CobingNo ratings yet
- Discussion Section of Paper For Biology - EditedDocument2 pagesDiscussion Section of Paper For Biology - EditedlucheNo ratings yet
- El Aprendizaje Aumenta La Supervivencia de Las Neuronas Recién Nacidas Siempre Que El Aprendizaje Sea Difícil de Lograr y Exitoso (2021)Document12 pagesEl Aprendizaje Aumenta La Supervivencia de Las Neuronas Recién Nacidas Siempre Que El Aprendizaje Sea Difícil de Lograr y Exitoso (2021)Catalina Muñoz MonteroNo ratings yet
- Tested Studies For Laboratory Teaching. Peifer. 1989Document174 pagesTested Studies For Laboratory Teaching. Peifer. 1989NidzaNo ratings yet
- Documenting Population Size and Survival Rate of Black-Capped ChickadeesDocument9 pagesDocumenting Population Size and Survival Rate of Black-Capped Chickadeesapi-338754159No ratings yet
- Question Answer SessionDocument2 pagesQuestion Answer Sessiontaner_soysurenNo ratings yet
- Stereotaxic Neurosurgery in Laboratory Rodent: Handbook on Best PracticesFrom EverandStereotaxic Neurosurgery in Laboratory Rodent: Handbook on Best PracticesNo ratings yet
- Battle of BritainDocument23 pagesBattle of BritainSimone MaucciNo ratings yet
- National Geographic History - JulyAugust 2023Document100 pagesNational Geographic History - JulyAugust 2023Renato LNo ratings yet
- Spatial-Numerical Associations: Shared Symbolic and Non-Symbolic Numerical RepresentationsDocument14 pagesSpatial-Numerical Associations: Shared Symbolic and Non-Symbolic Numerical RepresentationsSimone MaucciNo ratings yet
- Essay Writing GuideDocument25 pagesEssay Writing GuideChe HindsNo ratings yet
- 1 s2.0 S0149763422000896 MainDocument2 pages1 s2.0 S0149763422000896 MainSimone MaucciNo ratings yet
- Article - The Now-or-Never BottleneckDocument72 pagesArticle - The Now-or-Never BottleneckSimone MaucciNo ratings yet
- PreviewpdfDocument51 pagesPreviewpdfSimone MaucciNo ratings yet
- A Review of Time Management LiteratureDocument24 pagesA Review of Time Management LiteratureDrk AlbinusNo ratings yet
- Self-Control as Value-Based ChoiceDocument17 pagesSelf-Control as Value-Based ChoiceSimone MaucciNo ratings yet
- MSC ClinicalandTherapeuticNeuroscienceHandbook201920Document58 pagesMSC ClinicalandTherapeuticNeuroscienceHandbook201920Simone MaucciNo ratings yet
- Testolin McClelland-2020-Psychonomic Bulletin - ReviewDocument11 pagesTestolin McClelland-2020-Psychonomic Bulletin - ReviewSimone MaucciNo ratings yet
- Cerebral Lateralisation of First and Second LanguaDocument26 pagesCerebral Lateralisation of First and Second LanguaSimone MaucciNo ratings yet
- Personality and Politics: Values, Traits, and Political ChoiceDocument28 pagesPersonality and Politics: Values, Traits, and Political ChoiceSimone MaucciNo ratings yet
- Pnas 2203584119Document8 pagesPnas 2203584119Simone MaucciNo ratings yet
- 1 s2.0 S1059131115000382 MainDocument6 pages1 s2.0 S1059131115000382 MainSimone MaucciNo ratings yet
- S0028393221001354Document1 pageS0028393221001354Simone MaucciNo ratings yet
- 1 s2.0 S0028393221001354 MainDocument10 pages1 s2.0 S0028393221001354 MainSimone MaucciNo ratings yet
- How To Have Better Arguments With Your Romantic PartnerDocument1 pageHow To Have Better Arguments With Your Romantic PartnerSimone MaucciNo ratings yet
- Winter Variation - A Concise Guide to This Last Slot Cubing MethodDocument1 pageWinter Variation - A Concise Guide to This Last Slot Cubing MethodSimone Maucci100% (1)
- Introduction Bayesian InferenceDocument24 pagesIntroduction Bayesian InferenceSimone MaucciNo ratings yet
- Testolin - FrontHumNeurosc 2020Document9 pagesTestolin - FrontHumNeurosc 2020Simone MaucciNo ratings yet
- SpanishDocument2 pagesSpanishSimone MaucciNo ratings yet
- WVDocument2 pagesWVJoshua VictorNo ratings yet
- 50 Common English Phrasal Verbs PDFDocument54 pages50 Common English Phrasal Verbs PDFPapitch Do DuneNo ratings yet
- The Resolve ModelDocument10 pagesThe Resolve ModelSimone MaucciNo ratings yet
- The Resolve ModelDocument10 pagesThe Resolve ModelSimone MaucciNo ratings yet
- 5000 TOEFL Words PDFDocument36 pages5000 TOEFL Words PDFPrudhveeraj Chegu100% (2)
- SpanishDocument2 pagesSpanishSimone MaucciNo ratings yet
- Augoustinos, Walker - Social Cognition PDFDocument177 pagesAugoustinos, Walker - Social Cognition PDFSimone Maucci100% (2)
- Qognify-VisionHub-Brochure - Rev.01Document4 pagesQognify-VisionHub-Brochure - Rev.01Sarah BerlianiNo ratings yet
- 2016 CMCDocument536 pages2016 CMCStefan WeissNo ratings yet
- 2012 HCO Oriented Core ProceduresDocument30 pages2012 HCO Oriented Core ProceduresPancho Perez100% (1)
- 35.4 Uses Principles of Effective Speech Writing Focusing On Facial Expressions, Gestures and MovementsDocument4 pages35.4 Uses Principles of Effective Speech Writing Focusing On Facial Expressions, Gestures and MovementsWayne Dolorico MillamenaNo ratings yet
- F 09420010120134020 PP T 07Document19 pagesF 09420010120134020 PP T 07Stevia TjioeNo ratings yet
- Pellet PLNT ManualDocument19 pagesPellet PLNT ManualsubhankarprustyNo ratings yet
- Development of High Temperature Superconducting Transformers PDFDocument6 pagesDevelopment of High Temperature Superconducting Transformers PDFBen McConnellNo ratings yet
- University of Oxford, Financial Statements 2017-2018 PDFDocument120 pagesUniversity of Oxford, Financial Statements 2017-2018 PDFRano Digdayan MNo ratings yet
- Fabric in ArchitectureDocument28 pagesFabric in ArchitecturesubalakshmiNo ratings yet
- Linking Excel Data Into Navisworks - Part 5 Wrapping Up - RVITDocument2 pagesLinking Excel Data Into Navisworks - Part 5 Wrapping Up - RVITGreciaGuevaraNo ratings yet
- Bypass Sony KDL-55W900A 4X LED Blinking ErrorDocument7 pagesBypass Sony KDL-55W900A 4X LED Blinking ErroruzenNo ratings yet
- Aplac TR 001 Issue 2Document48 pagesAplac TR 001 Issue 2gaunananguyenNo ratings yet
- Error Correction ModelDocument37 pagesError Correction ModelRollins JohnNo ratings yet
- Diploma in Cyber Security - Data Communication and NetworkingDocument57 pagesDiploma in Cyber Security - Data Communication and Networkingsrikant.singh62583No ratings yet
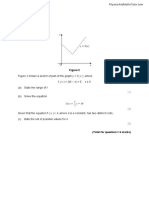
- Modulus of FunctionsDocument14 pagesModulus of FunctionsVajan SelvaratnamNo ratings yet
- 3-5 Support Document - Phase IDocument38 pages3-5 Support Document - Phase Iapi-290854496No ratings yet
- CSS - Proper Use and Maintenance of ToolsDocument1 pageCSS - Proper Use and Maintenance of ToolsRowell Marquina100% (1)
- Espedosa Module3.5 AssignmentDocument2 pagesEspedosa Module3.5 AssignmentKhemgee EspedosaNo ratings yet
- Selection and Characterisation of The Predominant Lactobacillus Species As A Starter Culture in The Preparation of Kocho, Fermented Food From EnsetDocument12 pagesSelection and Characterisation of The Predominant Lactobacillus Species As A Starter Culture in The Preparation of Kocho, Fermented Food From EnsetHelen WeldemichaelNo ratings yet
- RXF-C: Air Conditioning Technical DataDocument18 pagesRXF-C: Air Conditioning Technical DataJuan LezamaNo ratings yet
- Ds311 Specification Sheet EnglishDocument4 pagesDs311 Specification Sheet EnglishAntonio BocanegraNo ratings yet
- GL Poseidon User ManualDocument256 pagesGL Poseidon User ManualIrina MaltopolNo ratings yet
- Ophelia SyndromeDocument10 pagesOphelia SyndromeJulioroncal100% (1)
- Concepts - Essential Elements of Theories PDFDocument5 pagesConcepts - Essential Elements of Theories PDFKarlHeinrichNo ratings yet
- Sample Securitiser: Pressure Facility Re-InventedDocument4 pagesSample Securitiser: Pressure Facility Re-InventedMiguelNo ratings yet
- Saptamsa and Navamsa Charts PDFDocument37 pagesSaptamsa and Navamsa Charts PDFjyothiNo ratings yet
- Computer Networks: Humaira Ehsan Fall 2011Document18 pagesComputer Networks: Humaira Ehsan Fall 2011Bia JavedNo ratings yet
- Copper Pipe Leak Test and VacuumDocument2 pagesCopper Pipe Leak Test and Vacuumalone160162l100% (1)
- Waste Disposal Issues in Lalur VillageDocument27 pagesWaste Disposal Issues in Lalur VillageAnjana_EdathadanNo ratings yet
- Daily Stoic Amor Fati Excerpt From The Obstacle Is The WayDocument7 pagesDaily Stoic Amor Fati Excerpt From The Obstacle Is The WaySirLouen100% (9)