You might also like
- Acfrogdy87k6raqnhyzuaobv8dn5xllzhoemo34wgquexmrv3na6vzd6gxzbrvbnluowg F1nvq5qykblrx8r-Yewip3see Bwupqoyelggfn4gst7k252nphcdz7w9pgfhqchcomptkvk Kpc6qDocument11 pagesAcfrogdy87k6raqnhyzuaobv8dn5xllzhoemo34wgquexmrv3na6vzd6gxzbrvbnluowg F1nvq5qykblrx8r-Yewip3see Bwupqoyelggfn4gst7k252nphcdz7w9pgfhqchcomptkvk Kpc6qVerónica G-mNo ratings yet
- Amylase, Lipase, LDH, Trop I (Tabulated)Document9 pagesAmylase, Lipase, LDH, Trop I (Tabulated)maja.amora.swuNo ratings yet
- AutorDocument2 pagesAutorNuno CruzNo ratings yet
- Potential Role of Pro Renin Receptor in Cardiovascular and Kidney DiseasesDocument6 pagesPotential Role of Pro Renin Receptor in Cardiovascular and Kidney DiseasesAgata GavaNo ratings yet
- Vasoactive PeptidesDocument8 pagesVasoactive PeptidesSeff CausapinNo ratings yet
- Article Type Original ArticleDocument8 pagesArticle Type Original ArticleCarlos Alberto Martinez FloresNo ratings yet
- The Renin-Angiotensin Aldosterone System: Pathophysiological Role and Pharmacologic InhibitionDocument12 pagesThe Renin-Angiotensin Aldosterone System: Pathophysiological Role and Pharmacologic Inhibitionfatimah hasibuanNo ratings yet
- Therenin Angiotensinaldosteronesystem Final 120401080848 Phpapp02Document53 pagesTherenin Angiotensinaldosteronesystem Final 120401080848 Phpapp02andre kesumaNo ratings yet
- 2008 - Whole Genome PlatypusDocument11 pages2008 - Whole Genome Platypusj_wingerdNo ratings yet
- AsparaginaseDocument6 pagesAsparaginaseRegitaand PutNo ratings yet
- Mechanisms of Disease: Atriuretic EptidesDocument8 pagesMechanisms of Disease: Atriuretic EptidesВладимир ДружининNo ratings yet
- The Normal Kidney: Pediatrics 2 The Urinary System and Urinary Tract InfectionsDocument4 pagesThe Normal Kidney: Pediatrics 2 The Urinary System and Urinary Tract Infectionssarguss14No ratings yet
- Chicken or EggDocument1 pageChicken or Eggrajeshtalukdar420No ratings yet
- Unveiling The RNA World: Clinical Implications of Basic ResearchDocument3 pagesUnveiling The RNA World: Clinical Implications of Basic ResearchMauricio MerancioNo ratings yet
- ScientificDocument18 pagesScientificmatyastuma26No ratings yet
- Shinohara2016 PDFDocument8 pagesShinohara2016 PDFRafaelNo ratings yet
- 2023 - 01 - 23 1 - 17 PM Office LensDocument6 pages2023 - 01 - 23 1 - 17 PM Office LensKUMAR SUMITNo ratings yet
- Encyclopedia of Biological Chemistry - Vol - 3Document848 pagesEncyclopedia of Biological Chemistry - Vol - 3joshigauta100% (1)
- Nattokinase Prevents ThrombosesDocument3 pagesNattokinase Prevents ThrombosesmerrickNo ratings yet
- 1 s2.0 S1043276005000354 MainDocument6 pages1 s2.0 S1043276005000354 MaindasdNo ratings yet
- 01 Res 37 1 118Document10 pages01 Res 37 1 118KhanNo ratings yet
- James 1967 Anatomy of The Cardiac Conduction System in The RabbitDocument11 pagesJames 1967 Anatomy of The Cardiac Conduction System in The RabbitAna Katrina AguilaNo ratings yet
- Vasoactive PeptidesDocument8 pagesVasoactive PeptidesSeff CausapinNo ratings yet
- Immunological Cross-Reactivity Between Calcium ATPases From Human and Porcine ErythrocytesDocument7 pagesImmunological Cross-Reactivity Between Calcium ATPases From Human and Porcine ErythrocytesClement BewajiNo ratings yet
- Extrinsic Cardiac Conduction System AtfDocument2 pagesExtrinsic Cardiac Conduction System AtfMariaNo ratings yet
- 01 Hyp 9 1 41Document8 pages01 Hyp 9 1 41abdallaNo ratings yet
- Señalizacion RenalDocument62 pagesSeñalizacion Renalcesar eduardo castillejos cisnerosNo ratings yet
- Stahl 1976Document9 pagesStahl 1976Febriansyah Mohammed AkbarNo ratings yet
- Original Article: Rapamycin Combined with α-Cyanoacrylate Contributes to Inhibiting Intimal Hyperplasia in Rat ModelsDocument8 pagesOriginal Article: Rapamycin Combined with α-Cyanoacrylate Contributes to Inhibiting Intimal Hyperplasia in Rat ModelsMariantFomperosaNo ratings yet
- CP French Paradox 7janv10 enDocument2 pagesCP French Paradox 7janv10 enlelaissezfaireNo ratings yet
- Chronic Kidney Disease in Children New Trends in ManagementDocument103 pagesChronic Kidney Disease in Children New Trends in Managementfranklin ifioraNo ratings yet
- A Continuous Fluorescent Assay For The Determination of Plasma and Tissue Angiotensin I-Converting Enzyme ActivityDocument8 pagesA Continuous Fluorescent Assay For The Determination of Plasma and Tissue Angiotensin I-Converting Enzyme ActivityPablo GabrielNo ratings yet
- Early Detection of Ketoprofen-Induced Acute Kidney Injury in Sheep As Determined by Evaluation of Urinary Enzyme ActivitiesDocument7 pagesEarly Detection of Ketoprofen-Induced Acute Kidney Injury in Sheep As Determined by Evaluation of Urinary Enzyme ActivitiesCamilaGalvisNo ratings yet
- Enzyme Class E.C. Code Assay Reaction Tissue Source Clinical Significance Source of ErrorDocument3 pagesEnzyme Class E.C. Code Assay Reaction Tissue Source Clinical Significance Source of ErrorStephen YorNo ratings yet
- Cell Disease LabDocument1 pageCell Disease Labapi-340815123No ratings yet
- Acute PancreatitisDocument13 pagesAcute PancreatitisJacob BorongNo ratings yet
- Pepfidergic Innervafion of The Retinal Vasculature and Optic Nerve HeadDocument7 pagesPepfidergic Innervafion of The Retinal Vasculature and Optic Nerve HeadmoresubscriptionsNo ratings yet
- Increased Aspartate Aminotransferase Activity of Serum After in Vitro Supplementation With Pyridoxal PhosphateDocument7 pagesIncreased Aspartate Aminotransferase Activity of Serum After in Vitro Supplementation With Pyridoxal PhosphateAndi MusdalifahNo ratings yet
- PIIS0021925818615285Document6 pagesPIIS0021925818615285Lanceyy SanaminNo ratings yet
- Identification of Circular RNAs of Testis and Caput EpididymisDocument10 pagesIdentification of Circular RNAs of Testis and Caput EpididymisBahlibiNo ratings yet
- Basic Science Review: Functional Hyperemia in The BrainDocument7 pagesBasic Science Review: Functional Hyperemia in The BraindediNo ratings yet
- ACE Fix 2Document8 pagesACE Fix 2JantikaNo ratings yet
- Identification and Characteriization of Circular RNAs in Qinchuan Cattle TestisDocument10 pagesIdentification and Characteriization of Circular RNAs in Qinchuan Cattle TestisBahlibiNo ratings yet
- B-Mode and Doppler Ultrasound Imaging of The Spleen With Canine Splenic Torsion - A Retrospective EvaluationDocument5 pagesB-Mode and Doppler Ultrasound Imaging of The Spleen With Canine Splenic Torsion - A Retrospective EvaluationdenisegmeloNo ratings yet
- Clonorchiasis Manifesting As Obstructive Jaundice .1266 PDFDocument1 pageClonorchiasis Manifesting As Obstructive Jaundice .1266 PDFHighlander Emmanuel Fuentes BeltránNo ratings yet
- Enzymes CC Part 2 PrintDocument6 pagesEnzymes CC Part 2 PrintKrystel Bea DinqueNo ratings yet
- Argonaute PerspectiveDocument2 pagesArgonaute Perspectiveapi-3700537No ratings yet
- Effect of Isoxsuprine For Cervical Dilation in CowsDocument1 pageEffect of Isoxsuprine For Cervical Dilation in CowsGovind Narayan PurohitNo ratings yet
- Effect of IsoxsuprineDocument1 pageEffect of IsoxsuprinegnpobsNo ratings yet
- J. Biol. Chem. 1961 Rajagopalan 1059 65Document8 pagesJ. Biol. Chem. 1961 Rajagopalan 1059 65Cyrene MBolañosNo ratings yet
- Pnas 2118954119Document11 pagesPnas 2118954119FranekNo ratings yet
- 10 1021@acs Analchem 9b01570Document6 pages10 1021@acs Analchem 9b01570Iuliana SoldanescuNo ratings yet
- The Renin-Angiotensin System in The Year 2000: Review ArticleDocument18 pagesThe Renin-Angiotensin System in The Year 2000: Review ArticleEspinoza PaolaNo ratings yet
- 1987 - Epididymal Markers in Human InfertilityDocument11 pages1987 - Epididymal Markers in Human InfertilitymichelNo ratings yet
- Planos Avaliação PortalDocument4 pagesPlanos Avaliação PortalIzabela RodriguesNo ratings yet
- Diagnostic Procedure Day 3Document2 pagesDiagnostic Procedure Day 3Angeli RanocoNo ratings yet
- Lymphatic Drainage of The Neuraxis in Chronic Fatigue SyndromeDocument7 pagesLymphatic Drainage of The Neuraxis in Chronic Fatigue SyndromeAndrei CalabriaNo ratings yet
- Port Rna Integrity 2007Document12 pagesPort Rna Integrity 2007CeciliaPistolNo ratings yet
- Excretory SystemDocument5 pagesExcretory SystemveerdoriNo ratings yet
- Renin Angiotensin System and the HeartFrom EverandRenin Angiotensin System and the HeartWalmor C. De MelloNo ratings yet
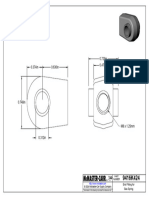
- 9416K424 - End Fitting For Gas SpringDocument1 page9416K424 - End Fitting For Gas SpringBaris YeltekinNo ratings yet
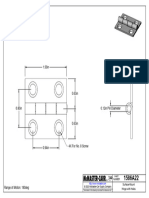
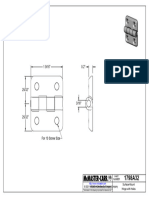
- 1586A22 - Surface-Mount Hinge With HolesDocument1 page1586A22 - Surface-Mount Hinge With HolesBaris YeltekinNo ratings yet
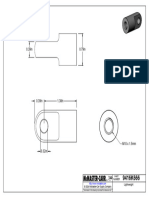
- 9416K666 LightweightDocument1 page9416K666 LightweightBaris YeltekinNo ratings yet
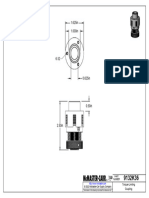
- 9132K19 - Torque-Limiting CouplingDocument1 page9132K19 - Torque-Limiting CouplingBaris YeltekinNo ratings yet
- 1798A32 - Surface-Mount Hinge With HolesDocument1 page1798A32 - Surface-Mount Hinge With HolesBaris YeltekinNo ratings yet
- 8356K13 - Electrical-Insulating Linear Sleeve BearingDocument1 page8356K13 - Electrical-Insulating Linear Sleeve BearingBaris YeltekinNo ratings yet
- Chain Hoist Dimensional DrawingDocument1 pageChain Hoist Dimensional DrawingBaris YeltekinNo ratings yet
- 2780T11 - High-Load Ball BearingDocument1 page2780T11 - High-Load Ball BearingBaris YeltekinNo ratings yet
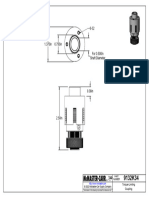
- 9132K36 - Torque-Limiting CouplingDocument1 page9132K36 - Torque-Limiting CouplingBaris YeltekinNo ratings yet
- 9132K34 - Torque-Limiting CouplingDocument1 page9132K34 - Torque-Limiting CouplingBaris YeltekinNo ratings yet
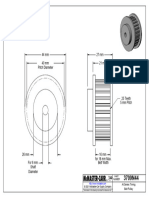
- 3709N44 - at Series Timing Belt PulleyDocument1 page3709N44 - at Series Timing Belt PulleyBaris YeltekinNo ratings yet
- Sensors: A Multiscale Region-Based Motion Detection and Background Subtraction AlgorithmDocument21 pagesSensors: A Multiscale Region-Based Motion Detection and Background Subtraction AlgorithmBaris YeltekinNo ratings yet
- Systems 195Document1 pageSystems 195Baris YeltekinNo ratings yet
- Hamilton Wheel W-2816-POYB-ULTRA-2Document1 pageHamilton Wheel W-2816-POYB-ULTRA-2Baris YeltekinNo ratings yet
- 7220K23 - Magnetic Torque-Limiting Shaft-to-Gear CouplingDocument1 page7220K23 - Magnetic Torque-Limiting Shaft-to-Gear CouplingBaris YeltekinNo ratings yet
- 9132K11 - Torque-Limiting CouplingDocument1 page9132K11 - Torque-Limiting CouplingBaris YeltekinNo ratings yet
- Dry LintDocument20 pagesDry LintBaris YeltekinNo ratings yet
- Iran Beyne 2 EnghelabDocument735 pagesIran Beyne 2 EnghelabBaris YeltekinNo ratings yet
- CM Electric Chain Hoists Specs and DrawingsDocument105 pagesCM Electric Chain Hoists Specs and DrawingsBaris YeltekinNo ratings yet
- Duct WrapDocument12 pagesDuct WrapBaris YeltekinNo ratings yet
- Welding Requirements Csa b3545 Masf Climbing Work PlatformsDocument2 pagesWelding Requirements Csa b3545 Masf Climbing Work PlatformsBaris YeltekinNo ratings yet
- Belt Conveyor Sizing ToolDocument4 pagesBelt Conveyor Sizing ToolBaris YeltekinNo ratings yet
- Series 1400Document9 pagesSeries 1400Baris YeltekinNo ratings yet
- SKF Seal MCW375-7000-312-H9BDocument3 pagesSKF Seal MCW375-7000-312-H9BBaris YeltekinNo ratings yet
- Hamilton Caster R EMD2 124FSTDocument1 pageHamilton Caster R EMD2 124FSTBaris YeltekinNo ratings yet
- An Introduction To Heavy Duty and Super Series ChainDocument15 pagesAn Introduction To Heavy Duty and Super Series ChainBaris YeltekinNo ratings yet
- UCFL 216: Flanged Ball Bearing Unit With Composite or Cast Metal HousingDocument5 pagesUCFL 216: Flanged Ball Bearing Unit With Composite or Cast Metal HousingBaris YeltekinNo ratings yet
- Predict Friction Lossin Slurry Pipes Darbyetal 1992Document5 pagesPredict Friction Lossin Slurry Pipes Darbyetal 1992Baris YeltekinNo ratings yet
- Autocentring of LensesDocument56 pagesAutocentring of LensesBaris YeltekinNo ratings yet
- Screw Jack CalculationDocument10 pagesScrew Jack CalculationBaris YeltekinNo ratings yet
- Unit 3 Chapter 9 DNA-Based Technologies LectureDocument20 pagesUnit 3 Chapter 9 DNA-Based Technologies LectureNeedBooksDontSueNo ratings yet
- Osteoporosis Edited 100201092008 Phpapp02Document32 pagesOsteoporosis Edited 100201092008 Phpapp02Maya SwariNo ratings yet
- 1184bns 1Document14 pages1184bns 1Daniela GanchasNo ratings yet
- Pathophysiology of Hypovolemic ShockDocument5 pagesPathophysiology of Hypovolemic ShockIan Rama100% (1)
- Mutaciones BGM PDFDocument6 pagesMutaciones BGM PDFwaldo tapiaNo ratings yet
- Experimental Eye ResearchDocument10 pagesExperimental Eye ResearchApriansyah Arfandy AzisNo ratings yet
- Stress-Induced (Takotsubo) CardiomyopathyDocument13 pagesStress-Induced (Takotsubo) CardiomyopathyAdrian GeambasuNo ratings yet
- Basic Principles of Real-Time PCRDocument19 pagesBasic Principles of Real-Time PCRMolecular_Diagnostics_KKUHNo ratings yet
- Biology Project: Lung CancerDocument12 pagesBiology Project: Lung CancerKarm ParmarNo ratings yet
- Prostaglandins Leukotrienes: Arthur Prancan, PH.DDocument34 pagesProstaglandins Leukotrienes: Arthur Prancan, PH.Dbahadar94No ratings yet
- PH, Titrable Acidity and Total Soluble Solids DeterminationDocument2 pagesPH, Titrable Acidity and Total Soluble Solids DeterminationcoffeecityNo ratings yet
- Homeostasis and Hormones AssignmentDocument4 pagesHomeostasis and Hormones AssignmentSivakumarran AgilanNo ratings yet
- Intrauterine Growth Retardation - IRA ONGDocument59 pagesIntrauterine Growth Retardation - IRA ONGaziz fahrudinNo ratings yet
- Why The NHS Must ChangeDocument2 pagesWhy The NHS Must ChangeMark HayesNo ratings yet
- The Red Cell Histogram PDFDocument9 pagesThe Red Cell Histogram PDFpieterinpretoria391No ratings yet
- 5 - DOWNLOAD Lactobacillus Acidophilus Rosell 52Document6 pages5 - DOWNLOAD Lactobacillus Acidophilus Rosell 52everstrongNo ratings yet
- Psychosocial Case Study FormatDocument3 pagesPsychosocial Case Study FormatLoh Wei Chieh0% (1)
- Nature ImmunologyDocument33 pagesNature ImmunologyололлNo ratings yet
- 08 - Microorganisms and Their Effects On Living ThingsDocument5 pages08 - Microorganisms and Their Effects On Living ThingsMie IsaNo ratings yet
- Hariri Et AlDocument4 pagesHariri Et AlJenniffer RiveraNo ratings yet
- Skin Barrier Function - Morphological Basis and Regulatory MechanismsDocument10 pagesSkin Barrier Function - Morphological Basis and Regulatory MechanismsJupiter CarlNo ratings yet
- Thesis ProposalDocument15 pagesThesis ProposalgkzunigaNo ratings yet
- CHE4171 - Revision (Final Exam)Document58 pagesCHE4171 - Revision (Final Exam)tiffanyyy00No ratings yet
- Stephanie Hall - Grief PowerpointDocument22 pagesStephanie Hall - Grief PowerpointricciardettoNo ratings yet
- Virulence Factors of Streptococcus PyogenesDocument6 pagesVirulence Factors of Streptococcus PyogenesAmador GielasNo ratings yet
- 12 Biology PP 23Document20 pages12 Biology PP 23Arsh ShrivastavNo ratings yet
- Swine in The Laboratory Surgery, Anesthesia, Imaging, and Experimental Techniques, Third Edition (Vetbooks - Ir)Document608 pagesSwine in The Laboratory Surgery, Anesthesia, Imaging, and Experimental Techniques, Third Edition (Vetbooks - Ir)marina stroeNo ratings yet
- PE - NovDec22 - Accommodating Multiple Modalities in The Same FacilityDocument8 pagesPE - NovDec22 - Accommodating Multiple Modalities in The Same FacilitysoceljoanNo ratings yet
- Thalassemias and HemoglobinopathiesDocument63 pagesThalassemias and HemoglobinopathiesMahmod_Al_Bust_4830No ratings yet
- Week 4.1 The Law of ResonanceDocument12 pagesWeek 4.1 The Law of ResonanceWim Massop100% (1)