You might also like
- INSECT PHYSIOLOGY AssignmentDocument12 pagesINSECT PHYSIOLOGY AssignmentLawal AbubakarNo ratings yet
- Digestive System: Food Uptake and UtilizationDocument12 pagesDigestive System: Food Uptake and UtilizationKarthi KarthiNo ratings yet
- Digestive System of BranchiostomaDocument6 pagesDigestive System of BranchiostomaAnuradha DasNo ratings yet
- Chinese General Hospital Colleges College of Nursing Anatomy and Physiology: Gastrointestinal System A. AnatomyDocument3 pagesChinese General Hospital Colleges College of Nursing Anatomy and Physiology: Gastrointestinal System A. AnatomyTin BernardezNo ratings yet
- AnaPhy - Digestive SystemDocument5 pagesAnaPhy - Digestive SystemJan Mark SotoNo ratings yet
- Digestive System - Lecture GuideDocument11 pagesDigestive System - Lecture GuideJEFFERSON ANDAYANo ratings yet
- Extra Reading Notes:: Name: Subject: Biology Class: X Date: 16 TO 25 MARCH, 2020 Topic: Life ProcessesDocument11 pagesExtra Reading Notes:: Name: Subject: Biology Class: X Date: 16 TO 25 MARCH, 2020 Topic: Life ProcessesHoney ChunduruNo ratings yet
- Unit Ten: Digestive SystemDocument9 pagesUnit Ten: Digestive SystemKath RubioNo ratings yet
- Digestive System HandoutsDocument7 pagesDigestive System HandoutsClexandrea Dela Luz Corpuz100% (1)
- Year 11 (WK 1-11) Biology Note (1st Term)Document62 pagesYear 11 (WK 1-11) Biology Note (1st Term)Keturah IbrahimNo ratings yet
- The Digestive SystemDocument23 pagesThe Digestive SystemJenderal Yohanes100% (1)
- Anatomy & Physiology - October 2-4Document15 pagesAnatomy & Physiology - October 2-4Ayessa Camelle DumileNo ratings yet
- Life ProcessesDocument28 pagesLife ProcessesVenkata SaiNo ratings yet
- Anatomy and Physiology For PudDocument6 pagesAnatomy and Physiology For PudBj Renquijo100% (1)
- Heterotrophic Nutrition PDFDocument10 pagesHeterotrophic Nutrition PDFNakshatra PaliwalNo ratings yet
- Life Processes Class 10 NotesDocument27 pagesLife Processes Class 10 NotesHarsh MauryaNo ratings yet
- PointersDocument5 pagesPointersKean AtonNo ratings yet
- Digestive System For Mono Gastric and Ruminant AnimalDocument9 pagesDigestive System For Mono Gastric and Ruminant AnimalAlex AdeyemiNo ratings yet
- Digestive System Anatomy and PhysiologyDocument18 pagesDigestive System Anatomy and PhysiologyKBDNo ratings yet
- Digestive System Anatomy PhysiologyDocument20 pagesDigestive System Anatomy PhysiologyKids JangNo ratings yet
- CHAPTER 28 - Animal Nutrition: The Need To Feed!Document24 pagesCHAPTER 28 - Animal Nutrition: The Need To Feed!Cathryne Kisha FloresNo ratings yet
- Anatomy ProjectDocument17 pagesAnatomy ProjectgilbertchenNo ratings yet
- Genbio2-Lesson 2-Nutrient Procurement and Processing Part 2Document45 pagesGenbio2-Lesson 2-Nutrient Procurement and Processing Part 2yvetteNo ratings yet
- Activity 9 Revised 1Document19 pagesActivity 9 Revised 1Jurist CoronelNo ratings yet
- Topic 8 Digestive SystemDocument10 pagesTopic 8 Digestive SystemMary Ann Morillo TenederoNo ratings yet
- Digestive System PartsDocument3 pagesDigestive System PartsDaniel DadzNo ratings yet
- Anatomy AND Physiology: (Gastrointestinal System)Document6 pagesAnatomy AND Physiology: (Gastrointestinal System)Antonette LeiNo ratings yet
- Frog Organ SystemDocument33 pagesFrog Organ SystemReineil PanguitoNo ratings yet
- Urinary Tract Infection Anatomy and PhysiologyDocument12 pagesUrinary Tract Infection Anatomy and PhysiologyPaul Anthony Centeno PimentelNo ratings yet
- Unit 8 The Digestive System: Learning OutcomesDocument12 pagesUnit 8 The Digestive System: Learning OutcomesDeolita BadiangNo ratings yet
- Nutrition in AnimalsDocument6 pagesNutrition in AnimalsRenuNo ratings yet
- DigestiveDocument5 pagesDigestivealayca cabatanaNo ratings yet
- Chapter I - Digestive SystemDocument3 pagesChapter I - Digestive SystemIndranil SinhaNo ratings yet
- Digestive SystemDocument2 pagesDigestive SystemQueencess Ara TorresNo ratings yet
- Insect NutritionDocument23 pagesInsect NutritionjobNo ratings yet
- Cholel FinalDocument39 pagesCholel FinalRizza DaleNo ratings yet
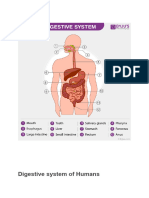
- Digestive System of HumansDocument13 pagesDigestive System of HumansNAMISH MAHAKULNo ratings yet
- Running Head: 1: The Process of Human DigestionDocument5 pagesRunning Head: 1: The Process of Human Digestiongmelendez2No ratings yet
- Reviewer ElsDocument8 pagesReviewer ElsAnsel Francis RapsingNo ratings yet
- BG3-3-The Digestive and Respiratory SystemsDocument36 pagesBG3-3-The Digestive and Respiratory Systemsapa@No ratings yet
- BOADO, Melody A. Lesson2 Act2 Clinical Application (NUPC106 Nutri..&Diet)Document4 pagesBOADO, Melody A. Lesson2 Act2 Clinical Application (NUPC106 Nutri..&Diet)Melody BoadoNo ratings yet
- Nutrition in Animals Class 7 Science Notes - Chapter 2: Different Ways of Taking FoodDocument4 pagesNutrition in Animals Class 7 Science Notes - Chapter 2: Different Ways of Taking FoodVikrant DeshmukhNo ratings yet
- Physiology of DigestionDocument92 pagesPhysiology of DigestionSiti Nur Lailatul MutmainnahNo ratings yet
- The Digestive System of Fish Consists ofDocument6 pagesThe Digestive System of Fish Consists ofPurity MoonNo ratings yet
- DIGESTIVEDocument6 pagesDIGESTIVEDanicaMae NinoNo ratings yet
- Digestive SystemDocument50 pagesDigestive SystemAlexander VonPatrickNo ratings yet
- Delhi Public School, Aligarh Biology Notes Chapter: Life Processes Teacher: Tanu Yadav DATE: 24.04.20 Day: FridayDocument29 pagesDelhi Public School, Aligarh Biology Notes Chapter: Life Processes Teacher: Tanu Yadav DATE: 24.04.20 Day: Fridayamrapali singhalNo ratings yet
- Digestive SystemDocument60 pagesDigestive SystemAnime tingzNo ratings yet
- Study of EarthwormDocument49 pagesStudy of EarthwormAshutosh RoutNo ratings yet
- Laboratorio de La Via Digestiva BajaDocument15 pagesLaboratorio de La Via Digestiva BajaDanna Lopez RamosNo ratings yet
- DIGESTIVE Lecture Notes PDFDocument20 pagesDIGESTIVE Lecture Notes PDFAaliya Minar100% (4)
- Karthik Bharadwaj'S: PresentationDocument32 pagesKarthik Bharadwaj'S: PresentationMahir AfranNo ratings yet
- The Human Digestive SystemDocument5 pagesThe Human Digestive SystemEuricka Preetty TañegaNo ratings yet
- TomdDocument15 pagesTomdduinnemerriedithcastroNo ratings yet
- E Pathshala File 801642381308Document48 pagesE Pathshala File 801642381308Aijaz AhmadNo ratings yet
- Functions of The Digestive SystemDocument8 pagesFunctions of The Digestive SystemPOTENCIANA MAROMANo ratings yet
- Anatomy and Physiology of Gastrointestinal TractDocument20 pagesAnatomy and Physiology of Gastrointestinal TractKrupa Jyothirmai100% (1)
- Stomach in VertebratesDocument6 pagesStomach in VertebratesTuhinNo ratings yet
- Presented By: Pramod Kumar MSC 1 Year RakconDocument37 pagesPresented By: Pramod Kumar MSC 1 Year Rakconpramod kumawatNo ratings yet
- Narrative Report-Day4Document3 pagesNarrative Report-Day4claire joy cabaseNo ratings yet
- Critique-Final CABASEDocument15 pagesCritique-Final CABASEclaire joy cabaseNo ratings yet
- Answer-Goat and Sheep Lab2Document7 pagesAnswer-Goat and Sheep Lab2claire joy cabaseNo ratings yet
- New Plant BreedingDocument4 pagesNew Plant Breedingclaire joy cabaseNo ratings yet
- Rules in OperationsDocument9 pagesRules in Operationsclaire joy cabaseNo ratings yet
- Human Reproductive System To BirthDocument11 pagesHuman Reproductive System To BirthDEEBANNo ratings yet
- Male and Female Reproductive SystemDocument4 pagesMale and Female Reproductive SystemJOSH ELORDENo ratings yet
- Human Anatomy Objectives WAPT 2022Document5 pagesHuman Anatomy Objectives WAPT 2022ExcelNo ratings yet
- Corrections and Additions To Taxonomists' Glossary of Mosquito AnatomyDocument17 pagesCorrections and Additions To Taxonomists' Glossary of Mosquito AnatomymokarahNo ratings yet
- Paper ReadingDocument3 pagesPaper ReadingRintan Novia Safitri0% (1)
- Endocrine System of FishesDocument5 pagesEndocrine System of FishesmonjitpaulNo ratings yet
- Animal Growth-Dev LPDocument3 pagesAnimal Growth-Dev LPTukta KanokpornNo ratings yet
- Raz lp36 Howanimalssleep CLRDocument9 pagesRaz lp36 Howanimalssleep CLRapi-290807931No ratings yet
- Why Is It Important To Learn About CellsDocument1 pageWhy Is It Important To Learn About Cellsrambo0% (1)
- Lesson Plan 2013 - TissueDocument25 pagesLesson Plan 2013 - Tissuesonyblank100% (1)
- Tugas Final Test EnglishDocument4 pagesTugas Final Test EnglishJiprayhanNo ratings yet
- Perpetuation of Life: Week 13 Earth & Life ScienceDocument23 pagesPerpetuation of Life: Week 13 Earth & Life ScienceREYNANTE SANTOSNo ratings yet
- L1 - Human BodyDocument40 pagesL1 - Human BodyFikril HakimNo ratings yet
- Arthropod ClassificationDocument6 pagesArthropod ClassificationFarzeen Mehmood100% (1)
- Virtual Fetal Pig DissectionDocument6 pagesVirtual Fetal Pig DissectionChristina DimitrijevicNo ratings yet
- Frog Dissection Answer SheetDocument4 pagesFrog Dissection Answer SheetAnde Falcone AlbiorNo ratings yet
- Human Body Systems: 7 Grade ScienceDocument17 pagesHuman Body Systems: 7 Grade Sciencenona wayne dela peñaNo ratings yet
- AleenaDocument1 pageAleenaanimefan26bNo ratings yet
- THOUGHT QUESTIONS (Second Exam)Document5 pagesTHOUGHT QUESTIONS (Second Exam)Lara Sabrina LumangNo ratings yet
- SkinDocument7 pagesSkincycyycNo ratings yet
- Introduction To General AnatomyDocument3 pagesIntroduction To General AnatomyNu Ri LeeNo ratings yet
- Cor 007 Module 17Document12 pagesCor 007 Module 17Camille ManlongatNo ratings yet
- Science9 q1 Mod1 SDOv2 1Document32 pagesScience9 q1 Mod1 SDOv2 1Amélie KatanaNo ratings yet
- Self CareDocument27 pagesSelf CareJessa May RapadasNo ratings yet
- Magician of The Sea - Reading Comprehension - StoryDocument2 pagesMagician of The Sea - Reading Comprehension - StoryHảo NguyễnNo ratings yet
- Laboratory Manual For Anatomy and Physiology 6th Edition Marieb Solutions ManualDocument26 pagesLaboratory Manual For Anatomy and Physiology 6th Edition Marieb Solutions ManualAlexMartinxfgr100% (56)
- GRADE 4 WEEK 4 and 5Document7 pagesGRADE 4 WEEK 4 and 5jommel vargasNo ratings yet
- Animal Kingdom PDFDocument2 pagesAnimal Kingdom PDFsyedzohairhusainrizviNo ratings yet
- Terminologia HistologicaDocument2 pagesTerminologia HistologicaLute CastroNo ratings yet
- Sensory Motor IntegrationDocument8 pagesSensory Motor IntegrationAdelinaPanaetNo ratings yet
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4 out of 5 stars4/5 (5)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (516)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (378)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- The Invention of Tomorrow: A Natural History of ForesightFrom EverandThe Invention of Tomorrow: A Natural History of ForesightRating: 4.5 out of 5 stars4.5/5 (5)
- Lymph & Longevity: The Untapped Secret to HealthFrom EverandLymph & Longevity: The Untapped Secret to HealthRating: 4.5 out of 5 stars4.5/5 (13)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (4)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- Change Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessFrom EverandChange Your Brain, Change Your Life (Before 25): Change Your Developing Mind for Real-World SuccessRating: 4 out of 5 stars4/5 (18)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemFrom EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemRating: 4.5 out of 5 stars4.5/5 (115)