You might also like
- AI and Blockchain in LawDocument11 pagesAI and Blockchain in LawIbrahim HassanNo ratings yet
- MIRPM Prospectus & Syllabus Autonomous-1Document30 pagesMIRPM Prospectus & Syllabus Autonomous-1Kanchan DewalNo ratings yet
- Company Law Course Outline 2023Document8 pagesCompany Law Course Outline 2023Louisa Hasford100% (1)
- Extent Managers Applied Inventory Control Approaches For Improving Operations of Small and Medium Scale Enterprises in South-East NigeriaDocument9 pagesExtent Managers Applied Inventory Control Approaches For Improving Operations of Small and Medium Scale Enterprises in South-East NigeriaInternational Journal of Recent Innovations in Academic ResearchNo ratings yet
- General Principles of Legal EthicsDocument6 pagesGeneral Principles of Legal EthicsAjaz GhaniNo ratings yet
- Primer: A Guide To The Organ-On - A - ChipDocument29 pagesPrimer: A Guide To The Organ-On - A - ChipAndreea CristinaNo ratings yet
- The AML Compliance Book - 150 Go - Matis MaekerDocument189 pagesThe AML Compliance Book - 150 Go - Matis Maekerbalup123No ratings yet
- 1.1.6 Lab - Cybersecurity Case Studies - ITExamAnswersc0baDocument2 pages1.1.6 Lab - Cybersecurity Case Studies - ITExamAnswersc0baabdel dalilNo ratings yet
- CHAPTER 4 Lesson 1 VirusDocument29 pagesCHAPTER 4 Lesson 1 VirusKreizel FajaNo ratings yet
- In Re Almacen, 31 SCRA 562 (1970)Document44 pagesIn Re Almacen, 31 SCRA 562 (1970)Joeleth Leoj BancayaNo ratings yet
- Methicillin-Resistant Staphylococcus Aureus PDFDocument23 pagesMethicillin-Resistant Staphylococcus Aureus PDFGerardo N. Pabón Gallino100% (1)
- A2 FormDocument5 pagesA2 Formshivam parmarNo ratings yet
- យុទ្ធសាស្ត្រការទូតសេដ្ឋកិច្ច ឆ្នាំ ២០២១-២០២៣ Economic Diplomancy Strategy 2021-2023Document26 pagesយុទ្ធសាស្ត្រការទូតសេដ្ឋកិច្ច ឆ្នាំ ២០២១-២០២៣ Economic Diplomancy Strategy 2021-2023Chan RithNo ratings yet
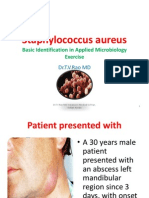
- Staphylococcus Aureus A Applied Microbiology ExerciseDocument27 pagesStaphylococcus Aureus A Applied Microbiology Exercisetummalapalli venkateswara raoNo ratings yet
- EC-Council Certified Incident Handler (ECIH) Course SummaryDocument7 pagesEC-Council Certified Incident Handler (ECIH) Course SummaryVinay KumarNo ratings yet
- Role of An Advocate in A Criminal Trial ProcessDocument5 pagesRole of An Advocate in A Criminal Trial ProcessDON ODUOR OTIENONo ratings yet
- Comparing LiFi and WiFi for Wireless ConnectivityDocument10 pagesComparing LiFi and WiFi for Wireless Connectivitystephenraj101 rajNo ratings yet
- 2023 Zscaler ThreatLabz Phishing ReportDocument39 pages2023 Zscaler ThreatLabz Phishing ReportSudhansu M NayakNo ratings yet
- Drafting of A LawDocument4 pagesDrafting of A LawIkram khanNo ratings yet
- Thigmotropism in Vigna Unguiculata Subsp. Sesquipedalis (C.N Sitaw)Document7 pagesThigmotropism in Vigna Unguiculata Subsp. Sesquipedalis (C.N Sitaw)Psychology and Education: A Multidisciplinary JournalNo ratings yet
- Bpo ProcessDocument4 pagesBpo ProcessAvick ChakrabortyNo ratings yet
- Exploring challenges facing Arabic ESL students and teachersDocument21 pagesExploring challenges facing Arabic ESL students and teachersMona English TutorNo ratings yet
- Trusted Digital Identity: BuildingDocument12 pagesTrusted Digital Identity: BuildingVítor CastroNo ratings yet
- Acronyms2 1Document17 pagesAcronyms2 1John SmithNo ratings yet
- GRI2009 EBrochureDocument27 pagesGRI2009 EBrochurenjaloualiNo ratings yet
- Liu2018 NAD Recommended by AttiaDocument20 pagesLiu2018 NAD Recommended by AttiaShantanu JhaNo ratings yet
- IoB for Improving Customer ServicesDocument4 pagesIoB for Improving Customer ServicesHannan GharadeNo ratings yet
- Lab Report 2Document5 pagesLab Report 2augustus willemNo ratings yet
- GI Tract InfectionsDocument29 pagesGI Tract InfectionsMahrukh SiddiquiNo ratings yet
- AG PROG Bachelor Master enDocument32 pagesAG PROG Bachelor Master enMargareta Fedora Prima ArtariNo ratings yet
- Legislative Drafting A New Sub-Discipline of Law Is BornDocument14 pagesLegislative Drafting A New Sub-Discipline of Law Is BornKhalid OsmanNo ratings yet
- D6587-12e1 Standard Test Method For Yarn Number Using Automatic TesterDocument6 pagesD6587-12e1 Standard Test Method For Yarn Number Using Automatic TesterAswindana Ibnu SenaNo ratings yet
- Forensics EvidenceDocument80 pagesForensics EvidenceSony BaniyaNo ratings yet
- A Review On - Data Hiding Using Cryptography and SteganographyDocument6 pagesA Review On - Data Hiding Using Cryptography and SteganographyiirNo ratings yet
- How To Prove ForgeryDocument35 pagesHow To Prove ForgeryAwkadigwe Fredrick IkennaNo ratings yet
- Catalogue Laboratory Products - Heraeus KulzerDocument288 pagesCatalogue Laboratory Products - Heraeus KulzerDentaDentNo ratings yet
- Hugh CollinsDocument10 pagesHugh CollinsBIPASHA KUNDUNo ratings yet
- Gender Sensitivity and Women's RightsDocument4 pagesGender Sensitivity and Women's RightsPjungNo ratings yet
- IPv4 Network Layer and AddressingDocument68 pagesIPv4 Network Layer and AddressingayeneNo ratings yet
- Practical ProgramsDocument9 pagesPractical Programshacking techNo ratings yet
- Model Contract of Apprenticeship Training For Major/Minor ApprenticesDocument3 pagesModel Contract of Apprenticeship Training For Major/Minor ApprenticesTejashree patilNo ratings yet
- Economics Project Final HimangDocument29 pagesEconomics Project Final HimangHimangSaraswatNo ratings yet
- Introduction to Wireless Sensor Networks (40 CharactersDocument79 pagesIntroduction to Wireless Sensor Networks (40 CharactersLoya SrijaNo ratings yet
- Scholarship Application FormDocument3 pagesScholarship Application Formchavawn mckenzieNo ratings yet
- Edelweiss Arbitrage Fund: Make Price Difference Between Markets Work For YouDocument16 pagesEdelweiss Arbitrage Fund: Make Price Difference Between Markets Work For YouNarendar KumarNo ratings yet
- 3- سياسة تغير الدخول الطوعي الي اجباريDocument4 pages3- سياسة تغير الدخول الطوعي الي اجباريralNo ratings yet
- AStudyonAnalysisofStockPricesofSelectedIndustries-research GateDocument51 pagesAStudyonAnalysisofStockPricesofSelectedIndustries-research GateAvicena Ilham GhifarieNo ratings yet
- Oracle Fusion Cloud SCM: Implementing Order ManagementDocument2,182 pagesOracle Fusion Cloud SCM: Implementing Order ManagementKannan RamanNo ratings yet
- 2002 RAeS Paper2751 V2Document28 pages2002 RAeS Paper2751 V2zishansinghNo ratings yet
- Ail TrainingDocument75 pagesAil TrainingSehivo100% (1)
- E-Governance CorruptionDocument23 pagesE-Governance CorruptionAnand SarvesvaranNo ratings yet
- Deep RL For BiologyDocument33 pagesDeep RL For BiologyMarcello BarylliNo ratings yet
- Nursing Concept Map TemplateDocument4 pagesNursing Concept Map TemplateAris Kendell BungabongNo ratings yet
- Intra-Uterine and Perinatal InfectionDocument72 pagesIntra-Uterine and Perinatal InfectionsabrinaNo ratings yet
- LogDocument13 pagesLog유리성100% (1)
- 80 Python Interview Questions & AnswersDocument23 pages80 Python Interview Questions & AnswersMahmoud ElhadyNo ratings yet
- Financial Risk Management & Derivatives: Unit 1Document30 pagesFinancial Risk Management & Derivatives: Unit 1DEEPIKA S R BUSINESS AND MANAGEMENT (BGR)No ratings yet
- CM V Alberta 2022 ABKB 716 DecisionDocument28 pagesCM V Alberta 2022 ABKB 716 DecisionAdam ToyNo ratings yet
- DNDR Guide 2nd EdDocument74 pagesDNDR Guide 2nd Edhaw lee100% (1)
- Prevalence and Antibiotic Resistance of Staphylococcus aureus in a Pakistani HospitalDocument4 pagesPrevalence and Antibiotic Resistance of Staphylococcus aureus in a Pakistani Hospitalzia ul RahmanNo ratings yet
- Ciaa 1717Document9 pagesCiaa 1717nada seddikiNo ratings yet
- 1 s2.0 S1074761314003987 MainDocument3 pages1 s2.0 S1074761314003987 Mainnada seddikiNo ratings yet
- Toxins 12 00080 v2Document22 pagesToxins 12 00080 v2nada seddikiNo ratings yet
- 45 Supplement - 3 S171Document6 pages45 Supplement - 3 S171nada seddikiNo ratings yet
- Hap Vap Guidelines 2016 PDFDocument51 pagesHap Vap Guidelines 2016 PDFJerryEddyaPutraBoerNo ratings yet
- Streptococcus Pneumoniae Staphylococci Faculty: Dr. Alvin FoxDocument32 pagesStreptococcus Pneumoniae Staphylococci Faculty: Dr. Alvin Foxdanish sultan100% (1)
- MULTIDRUG RESISTANCE MECHANISMSDocument14 pagesMULTIDRUG RESISTANCE MECHANISMSHarsh ChopraNo ratings yet
- GRAM Positive CocciDocument67 pagesGRAM Positive CocciNoraine Princess TabangcoraNo ratings yet
- Official Ncm0200 Baird Parker Agar Base Technical Specifications en UsDocument3 pagesOfficial Ncm0200 Baird Parker Agar Base Technical Specifications en UsBty SaGNo ratings yet
- 2010 Abstract Book PDFDocument109 pages2010 Abstract Book PDFManish MehraNo ratings yet
- Research Proposal For Tutuma NicholasDocument13 pagesResearch Proposal For Tutuma NicholasBit CoinNo ratings yet
- Novel Strategies For The Prevention and Treatment of Biofilm Related InfectionsDocument14 pagesNovel Strategies For The Prevention and Treatment of Biofilm Related Infectionsmonia agni wiyatamiNo ratings yet
- Paraben CompendiumDocument64 pagesParaben CompendiumAlma KunicNo ratings yet
- Steinberg 2009Document7 pagesSteinberg 2009majedNo ratings yet
- CMR 00047-19Document37 pagesCMR 00047-19UisanimgggNo ratings yet
- Recent Advances in Bacteriophage Therapy: How Delivery Routes, Formulation, Concentration and Timing Influence The Success of Phage TherapyDocument12 pagesRecent Advances in Bacteriophage Therapy: How Delivery Routes, Formulation, Concentration and Timing Influence The Success of Phage TherapyHammerly LinoNo ratings yet
- Phytochemical, antioxidant and antimicrobial analysis of selected mangrove speciesDocument6 pagesPhytochemical, antioxidant and antimicrobial analysis of selected mangrove speciesDanang RaharjoNo ratings yet
- MRSA FinalDocument4 pagesMRSA FinalApril DawnNo ratings yet
- 03 The Evaluation of Antibacterial Activity of Fabrics Impregnated With Dimethyltetradecyl (3 - (Trimethoxysilyl) Propyl) Ammonium ChlorideDocument8 pages03 The Evaluation of Antibacterial Activity of Fabrics Impregnated With Dimethyltetradecyl (3 - (Trimethoxysilyl) Propyl) Ammonium Chloridedharmayanti976No ratings yet
- DR Angelo Smith M.D WHPLDocument95 pagesDR Angelo Smith M.D WHPLMuhammad Riaz BhattiNo ratings yet
- Choosing the Right Antimicrobial AgentDocument53 pagesChoosing the Right Antimicrobial AgentEkanita DesianiNo ratings yet
- The Efficacy of Piper Betle Linn Against Methicillin Resistant Staphylococcus Aureus and Vancomycin-Resistant EnterococcusDocument20 pagesThe Efficacy of Piper Betle Linn Against Methicillin Resistant Staphylococcus Aureus and Vancomycin-Resistant EnterococcusClaire GonoNo ratings yet
- ICD-10-CM For Physicians 2018 Chapter Guidelines and Examples PDFDocument48 pagesICD-10-CM For Physicians 2018 Chapter Guidelines and Examples PDFRachelle AbuzoNo ratings yet
- Biological Exposure Control Plan Staphylococcus AureusDocument8 pagesBiological Exposure Control Plan Staphylococcus AureusferalianawasitoadhiNo ratings yet
- Antibiotics and Antibacterial DrugsDocument69 pagesAntibiotics and Antibacterial DrugsJames PerianayagamNo ratings yet
- Treatment Strategies For Methicillin-ResistantDocument12 pagesTreatment Strategies For Methicillin-ResistantSolRamirezNo ratings yet
- What's The Best Antibiotic For A Staph Infection - Treatments and RisksDocument3 pagesWhat's The Best Antibiotic For A Staph Infection - Treatments and RisksSundaramoorthy SelvanathanNo ratings yet
- Staphylococcus aureus Food PoisoningDocument9 pagesStaphylococcus aureus Food PoisoningAlifiana JatiningrumNo ratings yet
- Colony MorphologyDocument9 pagesColony MorphologyKirk AdderleyNo ratings yet
- Septic Arthritis in AdultsDocument19 pagesSeptic Arthritis in AdultsIvelisse TaverasNo ratings yet
- ThesisDocument30 pagesThesisjennifer bauzonNo ratings yet
- Bacterial InfectionsDocument47 pagesBacterial InfectionsSNo ratings yet
- European Journal of Pharmaceutics and BiopharmaceuticsDocument13 pagesEuropean Journal of Pharmaceutics and BiopharmaceuticsVareesshNo ratings yet
- Management of Diabetic Foot Ulcers: Pandji MulyonoDocument51 pagesManagement of Diabetic Foot Ulcers: Pandji MulyonoJeffrey Dyer100% (1)