You might also like
- G6PD Diet Foods To Avoid and IncludeDocument9 pagesG6PD Diet Foods To Avoid and IncludeJason Tiongco100% (6)
- RBC and Bleeding Disorders Outline Explains Anemia, Hemolytic DisordersDocument19 pagesRBC and Bleeding Disorders Outline Explains Anemia, Hemolytic DisordersNatan BabekNo ratings yet
- Note BiotechnologyDocument6 pagesNote BiotechnologyHala AlzuhairiNo ratings yet
- Carbohydrate Metabolism: By:-Dr - Priyanka Sharma 1 Year MDS Dept. of Public Health DentistryDocument93 pagesCarbohydrate Metabolism: By:-Dr - Priyanka Sharma 1 Year MDS Dept. of Public Health DentistrySimham Venu0% (1)
- Carbohydrate digestion and glucose metabolism pathwaysDocument4 pagesCarbohydrate digestion and glucose metabolism pathwaysJeremy EvansNo ratings yet
- 2Document38 pages2Kausik SenNo ratings yet
- 2023-2024-B3502-Lecture 17-Carbohydrate Metabolism-Gluconeogenesis and Oxidative Pentose Phosphate PathwayDocument15 pages2023-2024-B3502-Lecture 17-Carbohydrate Metabolism-Gluconeogenesis and Oxidative Pentose Phosphate Pathwaypitsolerato60No ratings yet
- Chapter Viii - Carbohydrates MechanismDocument34 pagesChapter Viii - Carbohydrates MechanismAngelo AngelesNo ratings yet
- Glycolysis 3MD (3)Document48 pagesGlycolysis 3MD (3)gostrider0093sNo ratings yet
- Carbohydratemetabolism 140214034339 Phpapp01 ConvertedDocument93 pagesCarbohydratemetabolism 140214034339 Phpapp01 Convertedyixecix709No ratings yet
- Carbohydrates (CHO)Document190 pagesCarbohydrates (CHO)Mustafa KhandgawiNo ratings yet
- Lec.6 PPP Pathway PDFDocument5 pagesLec.6 PPP Pathway PDFhawraaNo ratings yet
- Lec32 F08 HandoutDocument11 pagesLec32 F08 HandoutSarayu Ragavan NairNo ratings yet
- Pentose Phosphate PathwayDocument32 pagesPentose Phosphate PathwayAbdimalik AliNo ratings yet
- Lecture 8 NinaDocument87 pagesLecture 8 Ninaachraf rabadiNo ratings yet
- MLS 218 HMPDocument8 pagesMLS 218 HMPZainabNo ratings yet
- Define GlycolysisDocument3 pagesDefine GlycolysisjhanensuarezNo ratings yet
- Metabolic Pathway of Carbohydrate and GlycolysisDocument22 pagesMetabolic Pathway of Carbohydrate and GlycolysisDarshansinh MahidaNo ratings yet
- Carbohydrate Metabolism PathwaysDocument62 pagesCarbohydrate Metabolism PathwaysJHINUK DASGUPTANo ratings yet
- METABOLISM OF CARBOHYDRATESDocument62 pagesMETABOLISM OF CARBOHYDRATESmohaysin pirinoNo ratings yet
- (CO8) Chemistry of VisionDocument7 pages(CO8) Chemistry of VisionCardoge DalisayNo ratings yet
- 9 Lec. Biochemistry (4th)Document18 pages9 Lec. Biochemistry (4th)Doctor SonuNo ratings yet
- Lecture 10 Second PPP PDFDocument16 pagesLecture 10 Second PPP PDFMadani TawfeeqNo ratings yet
- Met of CHODocument43 pagesMet of CHOKiya AlemuNo ratings yet
- 3 Glikolisis 9Document17 pages3 Glikolisis 9Rafi Ahmad FarhanNo ratings yet
- Carbohydrate Metabolism 2022Document59 pagesCarbohydrate Metabolism 2022Catherine SilokaNo ratings yet
- Backwards ReasoningDocument40 pagesBackwards Reasoningharshit chaudharyNo ratings yet
- Metabolism & GlycolysisDocument3 pagesMetabolism & Glycolysisvanshkashp8272No ratings yet
- Biochem CombinedDocument758 pagesBiochem CombinedTheBoss 20No ratings yet
- Carbo Meta, Glycolysis, Krebs Cycle, GluconeoDocument80 pagesCarbo Meta, Glycolysis, Krebs Cycle, GluconeoMoses MutsikwiNo ratings yet
- C454 Lect3 ViewDocument66 pagesC454 Lect3 ViewAli Ali AliNo ratings yet
- What Is Metabolism?: Topic: Carbohydrate BiosynthesisDocument16 pagesWhat Is Metabolism?: Topic: Carbohydrate BiosynthesisNaiomiNo ratings yet
- Lecture 03. Carbohydrate Metabolism. Common Pathway, Regulation and PathologyDocument80 pagesLecture 03. Carbohydrate Metabolism. Common Pathway, Regulation and PathologyВіталій Михайлович НечипорукNo ratings yet
- Assignment For Blood - PaysonDocument25 pagesAssignment For Blood - PaysonRodrigo Joseph PaysonNo ratings yet
- Lintasan Pentosa FosfatDocument7 pagesLintasan Pentosa FosfatMUHAMMAD RIDONo ratings yet
- Carbohydrates Metabolism NotesDocument15 pagesCarbohydrates Metabolism NotesShaheenNo ratings yet
- Carbohydrate MetabolismDocument53 pagesCarbohydrate Metabolismjehram navalesNo ratings yet
- Carbohydrate MetabolismDocument8 pagesCarbohydrate MetabolismChaudhari Saroj100% (1)
- Hexose Monophosphate PathwayDocument19 pagesHexose Monophosphate Pathwayspdharanimaran001No ratings yet
- Carbohydrate Metabolism-1Document22 pagesCarbohydrate Metabolism-1Marwah100% (2)
- Carbohydrates MetabolismDocument33 pagesCarbohydrates MetabolismGungun FalguniNo ratings yet
- Pentose Phosphate PathwayDocument35 pagesPentose Phosphate PathwayRacheal MphandeNo ratings yet
- Carbo Meta, Glycolysis, Krebs Cycle, GluconeoDocument31 pagesCarbo Meta, Glycolysis, Krebs Cycle, GluconeoKellyNo ratings yet
- Biochemistry-Exam 2 Heinz SchwarzkopfDocument162 pagesBiochemistry-Exam 2 Heinz SchwarzkopfthomasNo ratings yet
- The Biomedical Importance of CarbohydratesDocument38 pagesThe Biomedical Importance of CarbohydratesAgnes Treyssia SandewaNo ratings yet
- CHAPTER 22 Gluconeogenesis, Glycogen Metabolism, and The Pentose Phosphate PathwayDocument12 pagesCHAPTER 22 Gluconeogenesis, Glycogen Metabolism, and The Pentose Phosphate Pathway楊畯凱No ratings yet
- Bioenergetics and Lipid MetabolismDocument34 pagesBioenergetics and Lipid MetabolismM Daffa Abhista ReviansyahNo ratings yet
- Biochemistry SidesDocument13 pagesBiochemistry SidesYousif KashatNo ratings yet
- Unit I I Carbohydrates MetabolismDocument33 pagesUnit I I Carbohydrates Metabolismseno adi100% (2)
- Naveen.K.L Natural ScienceDocument15 pagesNaveen.K.L Natural Sciencekripa v k nairNo ratings yet
- Carbohydrates MetabolismDocument6 pagesCarbohydrates MetabolismAisha ShahfiqueeNo ratings yet
- Unit I I Carbohydrates MetabolismDocument33 pagesUnit I I Carbohydrates MetabolismAna-Maria MargeanNo ratings yet
- CHE 311 Lecture 4 GlycolysisDocument28 pagesCHE 311 Lecture 4 Glycolysisisaac mwanzaNo ratings yet
- BIOCHEMISTRY QUESTIONSDocument5 pagesBIOCHEMISTRY QUESTIONSperssivesimangavwa2002No ratings yet
- Glycolysis and TCA CycleDocument38 pagesGlycolysis and TCA CycleAboubakar Moalim Mahad moh'dNo ratings yet
- GlycolysisDocument19 pagesGlycolysisatifzeaNo ratings yet
- (3.19) CarbosMetab (PPP, Other Sugars & GAGs)Document74 pages(3.19) CarbosMetab (PPP, Other Sugars & GAGs)pamelaNo ratings yet
- Kanzul Eman Biochemistry 1st SemesterDocument19 pagesKanzul Eman Biochemistry 1st SemesterMuhammad FarhanNo ratings yet
- Unit I I Carbohydrates MetabolismDocument33 pagesUnit I I Carbohydrates Metabolismaliyu belloNo ratings yet
- Carbohydrate MetabolismDocument72 pagesCarbohydrate MetabolismJeffson BalmoresNo ratings yet
- Chemistry of CarbohydratesDocument10 pagesChemistry of CarbohydratesYahya Daham Zafeer SakhrNo ratings yet
- Lipid Signaling and MetabolismFrom EverandLipid Signaling and MetabolismJames M. NtambiNo ratings yet
- University of Duhok College of Medicine Module of Metabolism Student Book 2021-2022Document213 pagesUniversity of Duhok College of Medicine Module of Metabolism Student Book 2021-2022QasmNo ratings yet
- Clinical Problem Solving Module Teaches Essential Doctor SkillsDocument87 pagesClinical Problem Solving Module Teaches Essential Doctor SkillsQasmNo ratings yet
- Cell Ultrastructure: Tissues of The Body ModuleDocument38 pagesCell Ultrastructure: Tissues of The Body ModuleQasmNo ratings yet
- Qualitative Test of Lipids Unsaturation Test For LipidsDocument12 pagesQualitative Test of Lipids Unsaturation Test For LipidsQasmNo ratings yet
- Lipid Profile: HDL & LDL CholesterolDocument6 pagesLipid Profile: HDL & LDL CholesterolQasmNo ratings yet
- Renal Function TestsDocument8 pagesRenal Function TestsQasmNo ratings yet
- General Urine Analysis GuideDocument24 pagesGeneral Urine Analysis GuideQasmNo ratings yet
- Clinical Biochemistry: Lipid ProfileDocument7 pagesClinical Biochemistry: Lipid ProfileQasmNo ratings yet
- 2 - SpectrophotometryDocument17 pages2 - SpectrophotometryQasmNo ratings yet
- 3 - Blood Correc.Document17 pages3 - Blood Correc.QasmNo ratings yet
- New Born ScreeningDocument30 pagesNew Born ScreeningJastine Miguel EGUIANo ratings yet
- Pseudomonas: Situs Inversus, Due ToDocument12 pagesPseudomonas: Situs Inversus, Due ToAmulya ThotambailuNo ratings yet
- Lecture # 7 Pentose Phosphate PathwayDocument44 pagesLecture # 7 Pentose Phosphate PathwayDesy AnggrainiNo ratings yet
- BiologyDocument27 pagesBiologysasiNo ratings yet
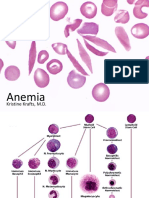
- Anemia: Kristine Krafts, M.DDocument38 pagesAnemia: Kristine Krafts, M.DNatalieMoralesNo ratings yet
- CH4 Patho D&R AgamDocument28 pagesCH4 Patho D&R AgamS Balagopal SivaprakasamNo ratings yet
- G6PD Deficiency Blood 1994Document24 pagesG6PD Deficiency Blood 1994Angela FurlanelloNo ratings yet
- G6PD DeficiencyDocument2 pagesG6PD DeficiencyShahylananda Tito YuwonoNo ratings yet
- Hemolytic AnemiaDocument61 pagesHemolytic AnemiaMangayarkarasi ChandrakasanNo ratings yet
- MCQ of HematologyDocument37 pagesMCQ of Hematologyalemante92% (13)
- The Child With Hematologic DisordersDocument149 pagesThe Child With Hematologic DisordersNics FranciscoNo ratings yet
- 1.1 Chromosomal Abnormalities: Hi H I Ef e FDocument26 pages1.1 Chromosomal Abnormalities: Hi H I Ef e FGlupiaSprawaNo ratings yet
- DH2289E G6PD EnglishDocument12 pagesDH2289E G6PD EnglishAmeer HamzaNo ratings yet
- Dr. Alfadel Alshaibani - Hematology Board ReviewDocument258 pagesDr. Alfadel Alshaibani - Hematology Board ReviewHanadi UmhanayNo ratings yet
- Inborn Errors of CHO MetabolismDocument30 pagesInborn Errors of CHO Metabolismwaleedemad649No ratings yet
- Pediatrics DR Masood Records NotesDocument24 pagesPediatrics DR Masood Records NotesM A100% (1)
- Biomarkers-A General ReviewDocument17 pagesBiomarkers-A General ReviewKevin FossNo ratings yet
- Formative Test 1 Week 1-2 Tropmed Odd 2023-2024 (Page 1 of 8) 2Document32 pagesFormative Test 1 Week 1-2 Tropmed Odd 2023-2024 (Page 1 of 8) 2Laura LatifaNo ratings yet
- Biochemistry QsDocument48 pagesBiochemistry QsFaisal AshiqNo ratings yet
- A Guide For G6PDDocument4 pagesA Guide For G6PDshimeath delrosarioNo ratings yet
- Guide To G6PD Deficiency Rapid Diagnostic Testing To Support Radical CureDocument34 pagesGuide To G6PD Deficiency Rapid Diagnostic Testing To Support Radical CureDadaNo ratings yet
- Anemia MonographDocument140 pagesAnemia MonographZaveri Hemant GirishkumarNo ratings yet
- Iron Deficiency Anemia GuideDocument78 pagesIron Deficiency Anemia GuidericaannpanugalinogNo ratings yet
- Chemical Examination of UrineDocument17 pagesChemical Examination of Urinekaran kambojNo ratings yet
- Acute Naphthalene Poisoning Following The Non-Accidental Ingestion of MothballsDocument4 pagesAcute Naphthalene Poisoning Following The Non-Accidental Ingestion of MothballsnaveenNo ratings yet
- G6PD Clinical DatabaseDocument8 pagesG6PD Clinical DatabaseCoy NuñezNo ratings yet
- Understanding and Managing Breast Milk JaundiceDocument7 pagesUnderstanding and Managing Breast Milk JaundiceJulian RiveraNo ratings yet
- Pentose Phosphate Pathway: M.Prasad Naidu MSC Medical Biochemistry, Ph.D.Research ScholarDocument22 pagesPentose Phosphate Pathway: M.Prasad Naidu MSC Medical Biochemistry, Ph.D.Research ScholarDr. M. Prasad NaiduNo ratings yet