You might also like
- The New Biology: A Battle between Mechanism and OrganicismFrom EverandThe New Biology: A Battle between Mechanism and OrganicismNo ratings yet
- The ontologies of organismic worldsDocument27 pagesThe ontologies of organismic worldsLamár ErzsébetNo ratings yet
- Stapleton e Froese - Is Collective Agency A Coherent IdeaDocument17 pagesStapleton e Froese - Is Collective Agency A Coherent IdeaPedro NoguezNo ratings yet
- Affect: Lisa Blackman and Couze VennDocument22 pagesAffect: Lisa Blackman and Couze VennaxiomatizadorrNo ratings yet
- FROESE, T. 2011 EnactiveApproachDocument36 pagesFROESE, T. 2011 EnactiveApproachFabiola CastilloNo ratings yet
- Sociality and The Life-Mind Continuity ThesisDocument25 pagesSociality and The Life-Mind Continuity ThesisandreanavarretehNo ratings yet
- Bapteste 2012Document26 pagesBapteste 2012raul roblesNo ratings yet
- Ontologies as Representations of RealityDocument12 pagesOntologies as Representations of Reality96325874145528725885No ratings yet
- Autopoiesis 40 Years LaterDocument25 pagesAutopoiesis 40 Years LaterRainman1618No ratings yet
- Autopoiesis, Adaptivity, Teleology, Agency: TEX PaperDocument18 pagesAutopoiesis, Adaptivity, Teleology, Agency: TEX PaperManuel Quezada CNo ratings yet
- Enactivism as a methodology in mathematics educationDocument7 pagesEnactivism as a methodology in mathematics educationDimitri MilleriNo ratings yet
- Autopoiesis 40 Years LaterDocument25 pagesAutopoiesis 40 Years LaterDavid Alcantara MirandaNo ratings yet
- The Circularity of The Embodied MindDocument13 pagesThe Circularity of The Embodied MindCoteOlivaPuginNo ratings yet
- Assessing The Prospects For A Return of PDFDocument32 pagesAssessing The Prospects For A Return of PDFManuel Quezada CNo ratings yet
- Being Embodied First-Person Judgements A PDFDocument9 pagesBeing Embodied First-Person Judgements A PDFkikaNo ratings yet
- Beyond Concepts: Ontology as Reality RepresentationDocument12 pagesBeyond Concepts: Ontology as Reality RepresentationdoctoratsrNo ratings yet
- Di Paolo, Ezequiel (2009) - "Overcoming Autopoiesis - An Enactive Detour On The Way From Life To SocietyDocument26 pagesDi Paolo, Ezequiel (2009) - "Overcoming Autopoiesis - An Enactive Detour On The Way From Life To SocietyDaniel Alva LeónNo ratings yet
- Enactive Cognitive Science Andbiology ofDocument2 pagesEnactive Cognitive Science Andbiology ofJosé Manuel Cruz OrtizNo ratings yet
- On The Logic of Animal Umwelten The AnimDocument14 pagesOn The Logic of Animal Umwelten The AnimMasterLoKing KingNo ratings yet
- The Apparent (Ur-)Intentionality of Living Beings and the Game of ContentDocument18 pagesThe Apparent (Ur-)Intentionality of Living Beings and the Game of ContentDeyvissonFernandesNo ratings yet
- The Broad Scope of EnactivismDocument2 pagesThe Broad Scope of EnactivismKe SaNo ratings yet
- Philosophy of Plant Morphology WorldviewsDocument4 pagesPhilosophy of Plant Morphology WorldviewsIvan RazoNo ratings yet
- Embodying The Mind and Representing The Body: Review of Philosophy and Psychology March 2012Document14 pagesEmbodying The Mind and Representing The Body: Review of Philosophy and Psychology March 2012biaNo ratings yet
- NESTEDDocument20 pagesNESTEDC CNo ratings yet
- 8 Coalitions - 1981 TurvyDocument73 pages8 Coalitions - 1981 TurvyjhoseffdouglasNo ratings yet
- Function and Functionalism, A Synthetic PerspectiveDocument21 pagesFunction and Functionalism, A Synthetic PerspectiveNicholas SmythNo ratings yet
- Function and Functionalism - Mahner2001Document20 pagesFunction and Functionalism - Mahner2001Emilia RaduNo ratings yet
- Steve Fleetwood: 1. An Ontology For Organisation and Management StudiesDocument40 pagesSteve Fleetwood: 1. An Ontology For Organisation and Management Studiesqwe rtyNo ratings yet
- Biosemiotics:: To Know, What Life KnowsDocument8 pagesBiosemiotics:: To Know, What Life Knowsbedevere86No ratings yet
- Embodied CognitionDocument8 pagesEmbodied Cognitionkashish behlNo ratings yet
- Hybrid Life: Integrating Biological, Artificial, and Cognitive SystemsDocument24 pagesHybrid Life: Integrating Biological, Artificial, and Cognitive SystemsOlaf WitkowskiNo ratings yet
- Phenomenological Teleology and Human IntDocument5 pagesPhenomenological Teleology and Human IntMasterLoKing KingNo ratings yet
- Gallagher 2018Document12 pagesGallagher 2018Rodrigo Toranzo (Roy)No ratings yet
- Gallagher, S., Making Enactivism Even More EmbodiedDocument16 pagesGallagher, S., Making Enactivism Even More Embodiedaxel16Hus13No ratings yet
- Crossing Over: The Interdisciplinary Meaning of Behavior Setting TheoryDocument10 pagesCrossing Over: The Interdisciplinary Meaning of Behavior Setting TheoryNurlia KhofiyyaNo ratings yet
- Organism Versus MechanismDocument8 pagesOrganism Versus MechanismdavidrojasvNo ratings yet
- Farnsworth 2017Document25 pagesFarnsworth 2017jbritoNo ratings yet
- The Function Debate in Philosophy ReviewedDocument29 pagesThe Function Debate in Philosophy Reviewedpde physicsNo ratings yet
- Module - CRS2Document22 pagesModule - CRS2Magr EscaNo ratings yet
- Enacting The World Seeking Clarity On en PDFDocument20 pagesEnacting The World Seeking Clarity On en PDFremistofelesNo ratings yet
- Viteritti 2013Document10 pagesViteritti 2013Hung DinhNo ratings yet
- Barrett, L. (2011) - Why Behaviorism Isn't Satanism (Cap. 2)Document22 pagesBarrett, L. (2011) - Why Behaviorism Isn't Satanism (Cap. 2)dana1520No ratings yet
- Accepted ManuscriptDocument26 pagesAccepted ManuscriptNavchetan RaiNo ratings yet
- Prospects For A Deep Phenomenology of EmbodimentDocument3 pagesProspects For A Deep Phenomenology of EmbodimentPatricia GallettiNo ratings yet
- Phenomenology and SomaestheticsDocument16 pagesPhenomenology and Somaesthetics曾慈鈞No ratings yet
- Autopoiesis, Biological Autonomy and The Process View of LifeDocument23 pagesAutopoiesis, Biological Autonomy and The Process View of LifeAlfonso Alatorre GarcíaNo ratings yet
- The Body As An Accumulation Strategy: Society and Space, 1995, 510)Document20 pagesThe Body As An Accumulation Strategy: Society and Space, 1995, 510)ganiesoktavianaNo ratings yet
- 6.GCTopoiOnlineFirstDocument14 pages6.GCTopoiOnlineFirstbryan zuñigaNo ratings yet
- Enactive Affectivity, ExtendedDocument11 pagesEnactive Affectivity, ExtendedTahi GuzmánNo ratings yet
- Behaviorism vs Cognitivism in Comparative Evolutionary PsychologyDocument23 pagesBehaviorism vs Cognitivism in Comparative Evolutionary PsychologyÁngel David Ayala OlmosNo ratings yet
- Making Sense of The Social Human Nonhuman Constellations and The Wicked Road To SustainabilityDocument11 pagesMaking Sense of The Social Human Nonhuman Constellations and The Wicked Road To SustainabilityDiana Carolina Hernandez PuentesNo ratings yet
- Making Sense of Sense-Making: Reflections On Enactive and Extended Mind TheoriesDocument8 pagesMaking Sense of Sense-Making: Reflections On Enactive and Extended Mind TheoriesanneoburgundaNo ratings yet
- 2012 The Body and The Experience of PresenceDocument17 pages2012 The Body and The Experience of PresenceCarlos A SanchesNo ratings yet
- A General Theory of EnactionDocument23 pagesA General Theory of EnactionBruce McNamaraNo ratings yet
- Critiquing Cartesian DualismDocument14 pagesCritiquing Cartesian DualismGeeKinqestNo ratings yet
- Carnal Sociology: Running HeadDocument7 pagesCarnal Sociology: Running HeadramshaNo ratings yet
- Behavioural Biologists Don't Agree On What Constitutes BehaviourDocument18 pagesBehavioural Biologists Don't Agree On What Constitutes BehaviourbuccaneerfosterNo ratings yet
- Allen - Teleological Notions in BiologyDocument15 pagesAllen - Teleological Notions in BiologyEma TakemuraNo ratings yet
- Self-Organization, Autonomy and The Bounds of The Self, Workhop 18-19 MarchDocument3 pagesSelf-Organization, Autonomy and The Bounds of The Self, Workhop 18-19 MarchandreaNo ratings yet
- Autopoiesis 40 Years Later 2012-Razeto-BarryDocument26 pagesAutopoiesis 40 Years Later 2012-Razeto-BarryIlse van der BeekNo ratings yet
- Consciousness From The Outside in and inDocument15 pagesConsciousness From The Outside in and inMasterLoKing KingNo ratings yet
- Dorfman - History of The Lifeworld. From Husserl To Merleau-PontyDocument10 pagesDorfman - History of The Lifeworld. From Husserl To Merleau-PontyAngelnecesarioNo ratings yet
- Perception As Body Body As Perception ReDocument17 pagesPerception As Body Body As Perception ReMasterLoKing KingNo ratings yet
- Invisibility From Discrimination To ResiDocument14 pagesInvisibility From Discrimination To ResiMasterLoKing KingNo ratings yet
- The Normative Turn in Enactive Theory AnDocument13 pagesThe Normative Turn in Enactive Theory AnMasterLoKing KingNo ratings yet
- Phenomenological Teleology and Human IntDocument5 pagesPhenomenological Teleology and Human IntMasterLoKing KingNo ratings yet
- Zavota FinalDocument31 pagesZavota FinalMasterLoKing KingNo ratings yet
- Hans Jonas and The Phenomenological Continuity of Life and MindDocument26 pagesHans Jonas and The Phenomenological Continuity of Life and MindMasterLoKing KingNo ratings yet
- Interpreting Saussure's Differential Theory: Merleau-Ponty Versus The LinguistsDocument42 pagesInterpreting Saussure's Differential Theory: Merleau-Ponty Versus The LinguistsMasterLoKing KingNo ratings yet
- Memes and The Exploitation of Imagination (Dennett, Daniel)Document10 pagesMemes and The Exploitation of Imagination (Dennett, Daniel)MasterLoKing KingNo ratings yet
- Legrand 2007Document17 pagesLegrand 2007MasterLoKing KingNo ratings yet
- The Bodily Self: The Sensori-Motor Roots of Pre-Reflective Self-ConsciousnessDocument30 pagesThe Bodily Self: The Sensori-Motor Roots of Pre-Reflective Self-ConsciousnessMasterLoKing KingNo ratings yet
- On The Logic of Animal Umwelten The AnimDocument14 pagesOn The Logic of Animal Umwelten The AnimMasterLoKing KingNo ratings yet
- Pre-Reflective Self-Consciousness On Being BodilyDocument28 pagesPre-Reflective Self-Consciousness On Being BodilyMasterLoKing KingNo ratings yet
- Syllabus of NCERT Booster Test Series - CoE NEET-2024 - 071157Document1 pageSyllabus of NCERT Booster Test Series - CoE NEET-2024 - 071157bishnuprasadmohapatra01No ratings yet
- 24160-Article Text-39228-1-10-20220603Document10 pages24160-Article Text-39228-1-10-20220603pandji pridjantoNo ratings yet
- Water Budget Equation Exercise Problems SolvedDocument10 pagesWater Budget Equation Exercise Problems SolvedSheeraz AhmedNo ratings yet
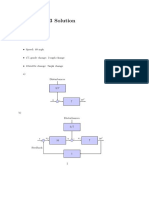
- Homework 3 Solution: Problem 1Document3 pagesHomework 3 Solution: Problem 1Gustavo Felicio PerruciNo ratings yet
- 10 Power Phrases To Master Your Next ArgumentDocument6 pages10 Power Phrases To Master Your Next ArgumentshaiannnicolebNo ratings yet
- This Study Resource Was: RSCH6352 Methods of ResearchDocument4 pagesThis Study Resource Was: RSCH6352 Methods of ResearchReuel Patrick CornagoNo ratings yet
- Lesson 4 Rainforest CharacteristicsDocument14 pagesLesson 4 Rainforest CharacteristicsLabib Kamal Year 11No ratings yet
- Seismic Retrofit in Hospitals Using Fluid ViscousDocument6 pagesSeismic Retrofit in Hospitals Using Fluid ViscousDeyanira Sayuri Yangali HerreraNo ratings yet
- The Polygamous Sex - Esther VilarDocument205 pagesThe Polygamous Sex - Esther VilarFederico Deambrosi Algorta100% (1)
- Actinometria QuimicaDocument42 pagesActinometria QuimicasgpizarroNo ratings yet
- Office PoliticsDocument17 pagesOffice Politicsnasif al islamNo ratings yet
- Math 5 - Pack 5Document42 pagesMath 5 - Pack 5Aneeza IftikharNo ratings yet
- Processing and Quality Evaluation of Guava SquashDocument3 pagesProcessing and Quality Evaluation of Guava SquashShailendra RajanNo ratings yet
- Evita Infinity v500 SW 2n Ifu 9052161 Ingles PDFDocument384 pagesEvita Infinity v500 SW 2n Ifu 9052161 Ingles PDFTu Papa2411No ratings yet
- Checklist For Welding and NDE SectionDocument2 pagesChecklist For Welding and NDE SectionMuhammadShabbirNo ratings yet
- Electric Forces, Fields, and Potential: LightningDocument164 pagesElectric Forces, Fields, and Potential: LightningIsmail Medhat SalahNo ratings yet
- Medical Assistant Essay ExamplesDocument2 pagesMedical Assistant Essay ExamplesbhhrxpaegNo ratings yet
- Understanding Schmitt TriggersDocument4 pagesUnderstanding Schmitt TriggersfernandofortunaNo ratings yet
- I've Got To Be MeDocument35 pagesI've Got To Be MeRosejen MangubatNo ratings yet
- Research 3Document76 pagesResearch 3Jackson IbascoNo ratings yet
- Modelling and simulating patient pathways at a Moroccan mother-child hospitalDocument7 pagesModelling and simulating patient pathways at a Moroccan mother-child hospitalashlyduartNo ratings yet
- Work EthicDocument57 pagesWork EthicRambo FranciscoNo ratings yet
- Lecture3 3Document27 pagesLecture3 3himan123No ratings yet
- kwb421 ManualDocument15 pageskwb421 ManualDante Obinu H.No ratings yet
- Feature Writing 2022Document51 pagesFeature Writing 2022lovelymae.madaliNo ratings yet
- Mixture: Learning SectionDocument19 pagesMixture: Learning SectionMd. Asif Islam UdayNo ratings yet
- Fundamentals of Research Methodology For Healthcare Professionals by Gisela Hildegard Van Rensburg Christa Van Der Walt Hilla BrinkDocument225 pagesFundamentals of Research Methodology For Healthcare Professionals by Gisela Hildegard Van Rensburg Christa Van Der Walt Hilla BrinkLeila RKNo ratings yet
- Climate Change and Conflict EssayDocument6 pagesClimate Change and Conflict EssayAgenyi AromeNo ratings yet
- (Form GNQ 15B) FrickCompPrestartChecklistDocument2 pages(Form GNQ 15B) FrickCompPrestartChecklisteugene mejidanaNo ratings yet
- UPP PE Tank Sump Installation GuideDocument32 pagesUPP PE Tank Sump Installation GuidelowiyaunNo ratings yet