You might also like
- Efecto de La Nutrición Sobre La Reproducción en Llamas y AlpacasDocument7 pagesEfecto de La Nutrición Sobre La Reproducción en Llamas y AlpacasALEX MOREANO ACOSTUPANo ratings yet
- Docchio 2019 Influence of Nutrition Body ConditiDocument8 pagesDocchio 2019 Influence of Nutrition Body ConditiAna SandovalNo ratings yet
- NutrientsDocument34 pagesNutrientssilvanaNo ratings yet
- Review Article: Integrative Control of Energy Balance and Reproduction in FemalesDocument14 pagesReview Article: Integrative Control of Energy Balance and Reproduction in FemalesfirdousNo ratings yet
- Nutrigenomics in Livestock - Recent AdvancesDocument11 pagesNutrigenomics in Livestock - Recent AdvancesIpung ArmstrongNo ratings yet
- 1 s2.0 S1751731114000731 MainDocument10 pages1 s2.0 S1751731114000731 MainEmmanuel CamargoNo ratings yet
- Impacts of Nutritional Management During Early Postnatal Life On Long-Term Physiological and Productive Responses of Beef CattleDocument11 pagesImpacts of Nutritional Management During Early Postnatal Life On Long-Term Physiological and Productive Responses of Beef CattleViníciusNo ratings yet
- Nutrition and Fertility in Ruminant Livestock 2006 RobinsonDocument18 pagesNutrition and Fertility in Ruminant Livestock 2006 RobinsonCanuto MunozNo ratings yet
- Nutrigenomics in Livestock and PoultryDocument19 pagesNutrigenomics in Livestock and PoultryRamachandran Ram86% (7)
- Voluntary Feed Intake in Growing-Finishing PigsDocument18 pagesVoluntary Feed Intake in Growing-Finishing PigsDiegoRodríguezSNo ratings yet
- Herdabilidade Bom!!Document18 pagesHerdabilidade Bom!!André MurotteNo ratings yet
- Applications of Nutrigenomics in Animal Science1Document24 pagesApplications of Nutrigenomics in Animal Science1Ramachandran RamNo ratings yet
- 1 s2.0 S1550413114000655 MainDocument13 pages1 s2.0 S1550413114000655 MainDesi AdestiNo ratings yet
- Multiovulación MoetDocument15 pagesMultiovulación MoetLuis GuachichulcaNo ratings yet
- Kayria Mapandi JunaidDocument9 pagesKayria Mapandi JunaidBarie ArnorolNo ratings yet
- AbstractDocument22 pagesAbstractnayayathNo ratings yet
- Physiology & BehaviorDocument9 pagesPhysiology & BehaviorBruno Garcia MontagniniNo ratings yet
- 1 s2.0 S1751731118001489 MainDocument15 pages1 s2.0 S1751731118001489 MainMaha RanuNo ratings yet
- The Future of Yogurt: Scientific and Regulatory NeedsDocument8 pagesThe Future of Yogurt: Scientific and Regulatory NeedsfredyNo ratings yet
- Modeling Dietary Influences On Offspring Metabolic Programming in Drosophila MelanogasterDocument12 pagesModeling Dietary Influences On Offspring Metabolic Programming in Drosophila MelanogasterDaveMartoneNo ratings yet
- Ajpendo 00495 2015Document18 pagesAjpendo 00495 2015siskaNo ratings yet
- Nutritional Impact On Gene Expression and Competence of Oocytes Used To Support Embryo Development and Livebirth by Cloning Procedures in GoatsDocument12 pagesNutritional Impact On Gene Expression and Competence of Oocytes Used To Support Embryo Development and Livebirth by Cloning Procedures in GoatsVerónica BecerraNo ratings yet
- Compensating For A Stressful Pregnancy Glucocorticoid Treatme - 2021 - HormonesDocument9 pagesCompensating For A Stressful Pregnancy Glucocorticoid Treatme - 2021 - HormonesVirginia-Mihaela Bruma2No ratings yet
- Pi Is 0022030220306287Document14 pagesPi Is 0022030220306287FranciscoNo ratings yet
- Combined Hormonal Contraceptives Are Associated With Minor Changes in Composition and Diversity in Gut Microbiota of Healthy WomenDocument11 pagesCombined Hormonal Contraceptives Are Associated With Minor Changes in Composition and Diversity in Gut Microbiota of Healthy Womensimon escobarNo ratings yet
- 2008 - QUIGLEY Et Al - Effect of Variable Long-Term Maternal Feed Allowance On The Development of The Ovine Placenta and FetusDocument10 pages2008 - QUIGLEY Et Al - Effect of Variable Long-Term Maternal Feed Allowance On The Development of The Ovine Placenta and FetusSamanta MonteiroNo ratings yet
- Dietary Intervention Prior To Pregnancy Reverses Metabolic Programming in Male Offspring of Obese RatsDocument9 pagesDietary Intervention Prior To Pregnancy Reverses Metabolic Programming in Male Offspring of Obese RatsKhaled Abdel-saterNo ratings yet
- Gut ResponseDocument17 pagesGut ResponseSeptinaAyuSamsiatiNo ratings yet
- Nutritional Requirements OverviewDocument6 pagesNutritional Requirements OverviewKoriaNo ratings yet
- Hormones and BehaviorDocument13 pagesHormones and BehaviorKerin ArdyNo ratings yet
- Masa TransisiDocument30 pagesMasa TransisiPrafangasti Sarah GinantikaNo ratings yet
- Science Reviewer 10Document3 pagesScience Reviewer 10Ivygrace NabitadNo ratings yet
- tmpE9EC TMPDocument9 pagestmpE9EC TMPFrontiersNo ratings yet
- Individual and Combined Effects of Anovulation and Cytological Endometritis On The Reproductive Performance of Dairy CowsDocument11 pagesIndividual and Combined Effects of Anovulation and Cytological Endometritis On The Reproductive Performance of Dairy CowsIoana CriveiNo ratings yet
- Article TodayDocument7 pagesArticle TodayGobardhan PudasainiNo ratings yet
- 1energy Balance and ReproductionDocument29 pages1energy Balance and ReproductionSterlin MorenoNo ratings yet
- Theriogenology: SciencedirectDocument6 pagesTheriogenology: SciencedirectGustav MolMedNo ratings yet
- SRD Innovative Technology Award 2010 Practical Aspects of The Fertility of Dairy CattleDocument17 pagesSRD Innovative Technology Award 2010 Practical Aspects of The Fertility of Dairy CattleIoana CriveiNo ratings yet
- A S F C S: Nimal Ourced Oods and Hild TuntingDocument18 pagesA S F C S: Nimal Ourced Oods and Hild Tuntingchandra dewiNo ratings yet
- CJAS09123 - Review Behaviour and Its Role in The Nutritional ManagementDocument9 pagesCJAS09123 - Review Behaviour and Its Role in The Nutritional Managementabdelaziz CHELIGHOUMNo ratings yet
- 1 s2.0 S1871141313000371 MainDocument6 pages1 s2.0 S1871141313000371 MainThiago F. MattosNo ratings yet
- Nutri GenomicsDocument9 pagesNutri GenomicsPromiseNo ratings yet
- Domestic Animal EndocrinologyDocument9 pagesDomestic Animal EndocrinologyJose Luis Ponce CovarrubiasNo ratings yet
- Script For Science ReportDocument3 pagesScript For Science Reportshn mzkNo ratings yet
- Article 3 PDFDocument10 pagesArticle 3 PDFSanae MoufidNo ratings yet
- The Role of Maternal Dietary Proteins in Development of Metabolic Syndrome in OffspringDocument33 pagesThe Role of Maternal Dietary Proteins in Development of Metabolic Syndrome in OffspringNurfadhilah YusufNo ratings yet
- Yang 2009Document18 pagesYang 2009Pamela DíazNo ratings yet
- Boland Et Al 2001 - Effect of Nutrition On Endocrine Parameters, Ovarian Physiology, and Oocyte and Embryo DevelopmentDocument18 pagesBoland Et Al 2001 - Effect of Nutrition On Endocrine Parameters, Ovarian Physiology, and Oocyte and Embryo DevelopmentsousamapNo ratings yet
- Jurnal 3Document15 pagesJurnal 3Prafangasti Sarah GinantikaNo ratings yet
- Nutritional Genomics: Patrick J. StoverDocument5 pagesNutritional Genomics: Patrick J. StoverTa KhanNo ratings yet
- Reproductive Systems Biology Tackles Global Issues of Population Growth, Food Safety and Reproductive HealthDocument5 pagesReproductive Systems Biology Tackles Global Issues of Population Growth, Food Safety and Reproductive HealthElla B. CollantesNo ratings yet
- A2 Option Module 8 Behaviour and Populations: Specification TableDocument18 pagesA2 Option Module 8 Behaviour and Populations: Specification Tablepunte77No ratings yet
- Global Food Security 23 (2019) 125-134Document10 pagesGlobal Food Security 23 (2019) 125-134fitria hidayantiNo ratings yet
- Nutrients 12 01734Document26 pagesNutrients 12 01734Arshy HzNo ratings yet
- Maternal Care PDFDocument27 pagesMaternal Care PDFAnonymous h4S1EeNo ratings yet
- Research in Veterinary Science: SciencedirectDocument9 pagesResearch in Veterinary Science: SciencedirectVen DedorNo ratings yet
- Progress in Research On Key Factors Regulating Lactation. Guo, H., Li, J., Wang, Y., Cao, X., LV, X., Yang, Z., & Chen, Z. (2023) .Document11 pagesProgress in Research On Key Factors Regulating Lactation. Guo, H., Li, J., Wang, Y., Cao, X., LV, X., Yang, Z., & Chen, Z. (2023) .Nxyz GxyzNo ratings yet
- Veterinary Medicine Sci - 2022 - Yi - Differences in Hormone Levels Around Parturition in Hanwoo Cattle Bos TaurusDocument6 pagesVeterinary Medicine Sci - 2022 - Yi - Differences in Hormone Levels Around Parturition in Hanwoo Cattle Bos TaurusAngélica CortésNo ratings yet
- International Workshop On Moringa and Nutrition: Accra, Ghana November 16-18, 2006Document37 pagesInternational Workshop On Moringa and Nutrition: Accra, Ghana November 16-18, 2006Farhat DurraniNo ratings yet
- Chesneau 2016Document7 pagesChesneau 2016Canuto MunozNo ratings yet
- Nutrition and Reproduction 1996 RobinsonDocument10 pagesNutrition and Reproduction 1996 RobinsonCanuto MunozNo ratings yet
- J ANIM SCI-2012-Fricke-pubertad en VaquillasDocument4 pagesJ ANIM SCI-2012-Fricke-pubertad en VaquillasCanuto MunozNo ratings yet
- Nutrition and Fertility in Ruminant Livestock 2006 RobinsonDocument18 pagesNutrition and Fertility in Ruminant Livestock 2006 RobinsonCanuto MunozNo ratings yet
- Live Weight and Metabolic Changes and The Associated Reproductive Performance in Maiden Ewes 2009 Ben SalemDocument5 pagesLive Weight and Metabolic Changes and The Associated Reproductive Performance in Maiden Ewes 2009 Ben SalemCanuto MunozNo ratings yet
- Kisspeptin Activates The Hypothalamic-Adenohypophyseal-Gonadal Axis in Prepubertal Ewe Lambs 2011 RedmondDocument8 pagesKisspeptin Activates The Hypothalamic-Adenohypophyseal-Gonadal Axis in Prepubertal Ewe Lambs 2011 RedmondCarlos MartínezNo ratings yet
- AGE at PUBERTY, FERTILITY and LITTER SIZE of Ewe Lambs Reared Under Different Photoregimens 1991 AinsworthDocument9 pagesAGE at PUBERTY, FERTILITY and LITTER SIZE of Ewe Lambs Reared Under Different Photoregimens 1991 AinsworthCanuto MunozNo ratings yet
- Dietary Protein or Energy Restriction Influences Age and Weight at Puberty in Ewes Lambs Boulanouar Et Al 1995Document10 pagesDietary Protein or Energy Restriction Influences Age and Weight at Puberty in Ewes Lambs Boulanouar Et Al 1995Canuto MunozNo ratings yet
- Age at Puberty and Fertility of Rahmani Sheep Fed On Biological Inoculated Corn Silage (2013) Khalifa y ColDocument10 pagesAge at Puberty and Fertility of Rahmani Sheep Fed On Biological Inoculated Corn Silage (2013) Khalifa y ColCanuto MunozNo ratings yet
- Age of Puberty in Ewes Which Developed Prenatally With Either A Ram or Ewe Fetus 1996 MeredithDocument4 pagesAge of Puberty in Ewes Which Developed Prenatally With Either A Ram or Ewe Fetus 1996 MeredithCanuto MunozNo ratings yet
- BMJ-Neonatal Jaundice PDFDocument54 pagesBMJ-Neonatal Jaundice PDFkhacthuanprovipNo ratings yet
- Obstetrics and GynaecologyDocument31 pagesObstetrics and GynaecologyLana LocoNo ratings yet
- Einc ScriptDocument3 pagesEinc ScriptJulia Rae Delos SantosNo ratings yet
- What are moral dilemmas? Understanding types and levelsDocument14 pagesWhat are moral dilemmas? Understanding types and levelsMeng lengNo ratings yet
- Mithun Rashi Baby Boy Names With MeaningsDocument7 pagesMithun Rashi Baby Boy Names With Meaningspoojamistry_123No ratings yet
- Lubchenco Curve PDF PDF Childhood Public HealthDocument1 pageLubchenco Curve PDF PDF Childhood Public HealthShaina MainitNo ratings yet
- Buhid-Mangyan Maternal HealthDocument11 pagesBuhid-Mangyan Maternal HealthQleoPsyche FadrigonNo ratings yet
- HIV SeminarDocument45 pagesHIV SeminarDelphy VargheseNo ratings yet
- CHAPTER TWO ANATOMY OF FEMALE PELVIS AND PhysiologyDocument62 pagesCHAPTER TWO ANATOMY OF FEMALE PELVIS AND PhysiologyabyNo ratings yet
- Obs1 41 Course Specification CorrectedDocument10 pagesObs1 41 Course Specification CorrectedWek KivNo ratings yet
- Utilization of Contraceptives Among Women of Reproductive Ages in Salaga MunicipalityDocument70 pagesUtilization of Contraceptives Among Women of Reproductive Ages in Salaga MunicipalitySeyram RichardNo ratings yet
- Notes-Chemical Coordination and Integration PDFDocument19 pagesNotes-Chemical Coordination and Integration PDFKisna guptaNo ratings yet
- Intrapartum Magnesium Sulfate Is Associated With Neuroprotection in Growth-Restricted FetusesDocument21 pagesIntrapartum Magnesium Sulfate Is Associated With Neuroprotection in Growth-Restricted FetusesHanifa YuniasariNo ratings yet
- Animal Reproduction MethodsDocument9 pagesAnimal Reproduction MethodsMelissa GerzonNo ratings yet
- Ovary Female Reproductive SystemDocument1 pageOvary Female Reproductive SystemtheodorastanerNo ratings yet
- BS Zoology: 5th Semester Placentation in MammalsDocument30 pagesBS Zoology: 5th Semester Placentation in MammalsTask BirdNo ratings yet
- Gynecological Examination Skill Repro Week 1 IndoDocument24 pagesGynecological Examination Skill Repro Week 1 Indowahyu kijang ramadhanNo ratings yet
- Hemingways Primitivism and Indian CampDocument13 pagesHemingways Primitivism and Indian Campzartaj rehamdinNo ratings yet
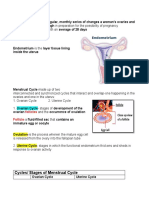
- Cycles/ Stages of Menstrual Cycle: Follicles FollicleDocument6 pagesCycles/ Stages of Menstrual Cycle: Follicles FollicleAaliyah CarlobosNo ratings yet
- Memorial For RespondentDocument34 pagesMemorial For RespondentmidhuNo ratings yet
- Journal of Health PromotionDocument2 pagesJournal of Health Promotiondevi ayu magviraNo ratings yet
- OB Return DemoDocument7 pagesOB Return DemoSophia RubiaNo ratings yet
- Science8 Q4 M3Document16 pagesScience8 Q4 M3Vine100% (1)
- Spontaneous Hepatic Rupture Associated With HELLP Syndrome: Britt C. Smyth, BA, RDMS, RVT, RDCSDocument3 pagesSpontaneous Hepatic Rupture Associated With HELLP Syndrome: Britt C. Smyth, BA, RDMS, RVT, RDCSMetharisaNo ratings yet
- Script - Agito Reniel A - BubbleplebDocument7 pagesScript - Agito Reniel A - BubbleplebReniel AgitoNo ratings yet
- Puberty, Sexuality, and Behavior: How To Guide Young People With DisabilitiesDocument55 pagesPuberty, Sexuality, and Behavior: How To Guide Young People With DisabilitiesDerison Marsinova100% (1)
- 8p23 Deletions FTNWDocument20 pages8p23 Deletions FTNWAdrian BoboceaNo ratings yet
- Prenatal DevelopmentDocument7 pagesPrenatal DevelopmentOlivia SnyderNo ratings yet
- Bio Chapter 47 Animal DevelopmentDocument15 pagesBio Chapter 47 Animal DevelopmentKAMAL KANT KUSHWAHANo ratings yet
- In Vitro Fertilization (Ivf)Document2 pagesIn Vitro Fertilization (Ivf)allfonsoNo ratings yet
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (378)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Inside of a Dog: What Dogs See, Smell, and KnowFrom EverandInside of a Dog: What Dogs See, Smell, and KnowRating: 4 out of 5 stars4/5 (390)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- Why We Sleep: Unlocking the Power of Sleep and DreamsFrom EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsRating: 4.5 out of 5 stars4.5/5 (2083)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (3)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- The Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsFrom EverandThe Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsNo ratings yet
- The Invention of Tomorrow: A Natural History of ForesightFrom EverandThe Invention of Tomorrow: A Natural History of ForesightRating: 4.5 out of 5 stars4.5/5 (5)