You might also like
- Aquaponics Systems, Fish. Volume 6: Sistemas de acuaponíaFrom EverandAquaponics Systems, Fish. Volume 6: Sistemas de acuaponíaNo ratings yet
- Acetic Acid FermentationDocument8 pagesAcetic Acid FermentationDan TanNo ratings yet
- Lactosuero 9 NoDocument24 pagesLactosuero 9 NoMarco Antonio Salcedo HinojosaNo ratings yet
- Optimised Production and Extraction of Astaxanthin From The Yeast Xanthophyllomyces DendrorhousDocument18 pagesOptimised Production and Extraction of Astaxanthin From The Yeast Xanthophyllomyces DendrorhousAndrea Alvarado RoNo ratings yet
- Bioprospecting of Exopolysaccharide-Producing Bacteria From Different Natural Ecosystems For Biopolymer Synthesis From VinasseDocument9 pagesBioprospecting of Exopolysaccharide-Producing Bacteria From Different Natural Ecosystems For Biopolymer Synthesis From VinasseVianney Casta¥Eda MonroyNo ratings yet
- 4 - Choubane S., Khelil O. Et Cheba B. 2015, African Journal of BiotechnologyDocument6 pages4 - Choubane S., Khelil O. Et Cheba B. 2015, African Journal of BiotechnologyOmar KHELILNo ratings yet
- Isolation and Characterization of Starter Culture From Spontaneous Fermentation of SourdoughDocument4 pagesIsolation and Characterization of Starter Culture From Spontaneous Fermentation of SourdoughHusna AdilaNo ratings yet
- End-Product Inhibition in Anaerobic Fermentations: Alejandro A. HerreroDocument5 pagesEnd-Product Inhibition in Anaerobic Fermentations: Alejandro A. Herreroalexnorton21No ratings yet
- Basis Protecting Oral Bacteria and An Enzymatic Role of The Arginine Deiminase System inDocument8 pagesBasis Protecting Oral Bacteria and An Enzymatic Role of The Arginine Deiminase System inAvishekh SinhaNo ratings yet
- Growth Studies of Potentially Probiotic Lactic Acid Bacteria in Cereal-Based SubstratesDocument9 pagesGrowth Studies of Potentially Probiotic Lactic Acid Bacteria in Cereal-Based SubstratesSjjshahaNo ratings yet
- Kadri 2018Document11 pagesKadri 2018Aslam MuhammadNo ratings yet
- Of Amaranthus Caudatus Seeds: in Vitro Antioxidant Effect and Inhibition ofDocument5 pagesOf Amaranthus Caudatus Seeds: in Vitro Antioxidant Effect and Inhibition ofJherson Gustavo SánchezNo ratings yet
- Eun-Ju Kim, Kwang-Moon Cho, Sun Jae Kwon, Seung-Ho Seo, Seong-Eun Park, Hong-Seok SonDocument8 pagesEun-Ju Kim, Kwang-Moon Cho, Sun Jae Kwon, Seung-Ho Seo, Seong-Eun Park, Hong-Seok SondianNo ratings yet
- Acido AceticoDocument17 pagesAcido AceticoOmar CanaveraNo ratings yet
- Bacillus Vallismortis TD6 (HQ992818)Document9 pagesBacillus Vallismortis TD6 (HQ992818)Karthikeyan SivashanmugamNo ratings yet
- Current Ethanol Production RequirementsDocument14 pagesCurrent Ethanol Production RequirementsSRINIVASAN TNo ratings yet
- Isolation and Identification of The Genera AcetobacterDocument9 pagesIsolation and Identification of The Genera AcetobacterervinamelaNo ratings yet
- VinegarDocument17 pagesVinegardeepikaNo ratings yet
- Biotechnology Progress - 2008 - Lenihan - Developing An Industrial Artemisinic Acid Fermentation Process To Support TheDocument7 pagesBiotechnology Progress - 2008 - Lenihan - Developing An Industrial Artemisinic Acid Fermentation Process To Support The774hzz9zd8No ratings yet
- Phenolic Compouns and Saponins in Quinoa Samples (Chenopodium Quinoa Willd.) Grown Under Different and Nonsaline Irrigation Regimens.Document8 pagesPhenolic Compouns and Saponins in Quinoa Samples (Chenopodium Quinoa Willd.) Grown Under Different and Nonsaline Irrigation Regimens.quiron333No ratings yet
- Rhizopus Oryzae: Production of L-Lactic Acid From Starch and Food Waste by Amylolytic MTCC 8784Document11 pagesRhizopus Oryzae: Production of L-Lactic Acid From Starch and Food Waste by Amylolytic MTCC 8784Christin SimamoraNo ratings yet
- Evaluation of The Alcoholic Fermentation Kinetics of Enzymatic Hydrolysates From Sugarcane Bagasse (Saccharum Officinarum L.)Document9 pagesEvaluation of The Alcoholic Fermentation Kinetics of Enzymatic Hydrolysates From Sugarcane Bagasse (Saccharum Officinarum L.)Felipe SouzaNo ratings yet
- Fermentation 04 00045Document12 pagesFermentation 04 00045hmossNo ratings yet
- 5.Applied-Acid Tolerant - KenjiTanakaDocument8 pages5.Applied-Acid Tolerant - KenjiTanakaImpact JournalsNo ratings yet
- Applied and Environmental Microbiology 1997 Narendranath 4158.fullDocument6 pagesApplied and Environmental Microbiology 1997 Narendranath 4158.fullKannan RanganathanNo ratings yet
- Dragon Fruit PDFDocument8 pagesDragon Fruit PDFNguyễn Võ Hoài PhươngNo ratings yet
- Aem00154 0187Document8 pagesAem00154 0187HonganhNo ratings yet
- Summary of Bioethanol Production From Sargassum Spp.Document1 pageSummary of Bioethanol Production From Sargassum Spp.Agil Ramadhan PNo ratings yet
- Role of The Arginine Deiminase System Protecting Enzymatic ToleranceDocument7 pagesRole of The Arginine Deiminase System Protecting Enzymatic ToleranceAvishekh SinhaNo ratings yet
- Happiemaga2008 PDFDocument9 pagesHappiemaga2008 PDFGabriel FigueroaNo ratings yet
- Bioethanol Et Production From Mango YeasteDocument5 pagesBioethanol Et Production From Mango YeasteNauden Maïmouna SILLUENo ratings yet
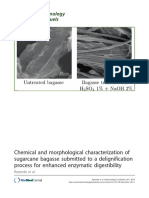
- Chemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityDocument19 pagesChemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityTabiita Huaman NorabuenaNo ratings yet
- 11004-Article Text PDF-31187-2-10-20180516Document12 pages11004-Article Text PDF-31187-2-10-20180516BrianWirawanNo ratings yet
- International Journal of Green and Herbal ChemistryDocument8 pagesInternational Journal of Green and Herbal ChemistryMat NeriNo ratings yet
- Journal PaperDocument8 pagesJournal PaperraguthiyaguNo ratings yet
- MesophilDocument4 pagesMesophilnurcincelikNo ratings yet
- Renewable Energy: Tesfayesus Zinare Mamo, Abhishek Dutta, S. Anuradha JabasinghDocument6 pagesRenewable Energy: Tesfayesus Zinare Mamo, Abhishek Dutta, S. Anuradha JabasinghSatawat TanaratNo ratings yet
- A Thermotolerant and High Acetic Acid-Producing Bacterium Acetobacter SPDocument8 pagesA Thermotolerant and High Acetic Acid-Producing Bacterium Acetobacter SPWendy RiveraNo ratings yet
- Acid Acetic Removal Vinuri LevuriDocument4 pagesAcid Acetic Removal Vinuri LevuriRazvan FilimonNo ratings yet
- Lipase 3Document9 pagesLipase 3Angie Grados LopezNo ratings yet
- Ramos-Suárez and Carreras (2014) - Use of Microalgae Residues For Biogas Production PDFDocument10 pagesRamos-Suárez and Carreras (2014) - Use of Microalgae Residues For Biogas Production PDFZouzo222No ratings yet
- Zosterisessor Ophiocephalus, Raja Clavata Scorpaena Scrofa: Research ArticleDocument10 pagesZosterisessor Ophiocephalus, Raja Clavata Scorpaena Scrofa: Research ArticleLaila Nurul QodryNo ratings yet
- Biomass and Bioenergy: Research PaperDocument10 pagesBiomass and Bioenergy: Research PaperPaulina Lpz JrzNo ratings yet
- Anaerobic Thermophilic Fermentation For Acetic Acid Production From Milk PermeateDocument10 pagesAnaerobic Thermophilic Fermentation For Acetic Acid Production From Milk PermeateViet_Len_Troi_XanhNo ratings yet
- Answers To The Questions of Chapter 1Document14 pagesAnswers To The Questions of Chapter 1kellyleeeeNo ratings yet
- Backup of ACETIC ACID - FinalDocument95 pagesBackup of ACETIC ACID - FinalMontarum, Kathleen R.No ratings yet
- Montoya-Rodríguez Alvaro, 2012. Publicado en AMDocument10 pagesMontoya-Rodríguez Alvaro, 2012. Publicado en AMRoberto GutiérrezNo ratings yet
- Optimization of Process Parameters Salinity, Temperature and PH For The Growth of Sp. S 27Document25 pagesOptimization of Process Parameters Salinity, Temperature and PH For The Growth of Sp. S 27Allyssa Jiselle CabalongaNo ratings yet
- Fermentation of Lactose To Ethanol in Cheese WheyDocument13 pagesFermentation of Lactose To Ethanol in Cheese WheyVitor ArsenioNo ratings yet
- Fermentation of Lactose To Ethanol in Cheese Whey Permeate and Concentrated Permeate by Engineered Escherichia ColiDocument12 pagesFermentation of Lactose To Ethanol in Cheese Whey Permeate and Concentrated Permeate by Engineered Escherichia ColijoelfloresanchezNo ratings yet
- Optimization of Kojic Acid Production Conditions From Cane Molasses Using Plackett-Burman DesignDocument14 pagesOptimization of Kojic Acid Production Conditions From Cane Molasses Using Plackett-Burman DesignFaelFernandesNo ratings yet
- KarotenoidDocument9 pagesKarotenoidChristina Dwi KusumaningtyasNo ratings yet
- En Zima Conserva Ça oDocument7 pagesEn Zima Conserva Ça oJonas TeixeiraNo ratings yet
- Acido LacticoDocument9 pagesAcido LacticoJuan Victor ChampeNo ratings yet
- Interactions Between Saccharomyces Cerevisiae and Lactic Acid Bacteria in SourdoughDocument5 pagesInteractions Between Saccharomyces Cerevisiae and Lactic Acid Bacteria in SourdoughmaurodiloretoNo ratings yet
- DONAJIDocument8 pagesDONAJImartinNo ratings yet
- 2011 Basheer Et Al Aspergillus Awamori Lipase ProductionDocument12 pages2011 Basheer Et Al Aspergillus Awamori Lipase ProductiontigapeptindonesiaNo ratings yet
- 1 s2.0 S1570023211007112 MainDocument6 pages1 s2.0 S1570023211007112 MainyasserNo ratings yet
- Minitab17 GettingStarted enDocument6 pagesMinitab17 GettingStarted enTan Hau VoNo ratings yet
- Research Article Bacillus SubtilisDocument10 pagesResearch Article Bacillus SubtilisDương Nguyễn Thùy DungNo ratings yet
- Bacterial MetabolismDocument13 pagesBacterial MetabolismololadeNo ratings yet
- Introductory MicroBiology With PracticalsDocument118 pagesIntroductory MicroBiology With Practicalsmatrixxxx420100% (1)
- Vocabulary WordsDocument2 pagesVocabulary WordsLennie DiazNo ratings yet
- Data in BriefDocument6 pagesData in BriefMuhammad aldi sofyanNo ratings yet
- Mass BalanceDocument45 pagesMass BalanceDafik A. MasruriNo ratings yet
- Microbial Metabolism-The Chemical Crossroads of LifeDocument73 pagesMicrobial Metabolism-The Chemical Crossroads of Lifepranky neyneyNo ratings yet
- Yeast RespirationDocument5 pagesYeast RespirationPauline LapuzNo ratings yet
- ContentServer PDFDocument10 pagesContentServer PDFSilvia Patricia Rodríguez Peña 2171033No ratings yet
- AP Lab #6 Cellular RespirationDocument7 pagesAP Lab #6 Cellular RespirationAnya KureshiNo ratings yet
- Renewable and Sustainable Energy ReviewsDocument30 pagesRenewable and Sustainable Energy ReviewsMARIA CAMILA MUÑOZ VEGANo ratings yet
- A2 Biology Syllabus 9700: Unit 1: Energy, Respiration and PhotosynthesisDocument18 pagesA2 Biology Syllabus 9700: Unit 1: Energy, Respiration and PhotosynthesisaduhaikNo ratings yet
- Commercial Applications of EnzymesDocument3 pagesCommercial Applications of EnzymesJudy Ann BoseNo ratings yet
- Cell Respiration FermentationDocument5 pagesCell Respiration FermentationRozuwan ZakariaNo ratings yet
- Microbial Products of Potential Importance - Product / Activity Examples ProductsDocument10 pagesMicrobial Products of Potential Importance - Product / Activity Examples ProductsShveta Jaishankar Iyer100% (4)
- Yeast ExperimentDocument2 pagesYeast ExperimentJames Knowell0% (1)
- Detailed Project Report (DPR) On 100 KLPD Multifeed Distillery Plant Based On Molasses and GrainsDocument81 pagesDetailed Project Report (DPR) On 100 KLPD Multifeed Distillery Plant Based On Molasses and GrainsNitipal SinghNo ratings yet
- Food SpoilageDocument42 pagesFood Spoilagedrgunavet100% (6)
- Automated Control of Cocoa Post-Harvest and Transformation Process To Obtain High-Quality BeansDocument6 pagesAutomated Control of Cocoa Post-Harvest and Transformation Process To Obtain High-Quality Beanscacalot93No ratings yet
- Sithccc041: Produce Cakes, Pastries, and BreadDocument45 pagesSithccc041: Produce Cakes, Pastries, and Breadryanmai2804No ratings yet
- Wasseraufbereitung enDocument16 pagesWasseraufbereitung enVera VeroniqueNo ratings yet
- Advantages and Disadvantages of Traditional Fermentation of Dairy ProductsDocument4 pagesAdvantages and Disadvantages of Traditional Fermentation of Dairy ProductsEkoh EnduranceNo ratings yet
- CancerDocument15 pagesCancerGordo y TutucaNo ratings yet
- Module-9 (Theory) Media FormulationDocument4 pagesModule-9 (Theory) Media FormulationriyaNo ratings yet
- The Chemistry of Cooking and Cleaning - Ellen RichardsDocument106 pagesThe Chemistry of Cooking and Cleaning - Ellen RichardsSandra Mian100% (2)
- Experiment 3 Bio 150Document4 pagesExperiment 3 Bio 150nordiana muhidinNo ratings yet
- Butanol & EthanolDocument17 pagesButanol & EthanolnivethaNo ratings yet
- MetabolismeDocument31 pagesMetabolismeLalu Aldi PratamaNo ratings yet
- Desta AlcoholDocument76 pagesDesta AlcoholMULAT DEGAREGENo ratings yet
- Establishment of A Industrial ProcessDocument12 pagesEstablishment of A Industrial ProcessIbrahimAslamNo ratings yet
- Coconut Oil Production &refiningDocument5 pagesCoconut Oil Production &refiningAmylene Corral AliganNo ratings yet
- Is That a Fact?: Frauds, Quacks, and the Real Science of Everyday LifeFrom EverandIs That a Fact?: Frauds, Quacks, and the Real Science of Everyday LifeRating: 4.5 out of 5 stars4.5/5 (3)
- Meltdown: Nuclear disaster and the human cost of going criticalFrom EverandMeltdown: Nuclear disaster and the human cost of going criticalRating: 5 out of 5 stars5/5 (5)
- The Elements We Live By: How Iron Helps Us Breathe, Potassium Lets Us See, and Other Surprising Superpowers of the Periodic TableFrom EverandThe Elements We Live By: How Iron Helps Us Breathe, Potassium Lets Us See, and Other Surprising Superpowers of the Periodic TableRating: 3.5 out of 5 stars3.5/5 (22)
- Chemistry: a QuickStudy Laminated Reference GuideFrom EverandChemistry: a QuickStudy Laminated Reference GuideRating: 5 out of 5 stars5/5 (1)
- Organic Chemistry for Schools: Advanced Level and Senior High SchoolFrom EverandOrganic Chemistry for Schools: Advanced Level and Senior High SchoolNo ratings yet
- Periodic Tales: A Cultural History of the Elements, from Arsenic to ZincFrom EverandPeriodic Tales: A Cultural History of the Elements, from Arsenic to ZincRating: 3.5 out of 5 stars3.5/5 (137)
- The Disappearing Spoon: And Other True Tales of Madness, Love, and the History of the World from the Periodic Table of the ElementsFrom EverandThe Disappearing Spoon: And Other True Tales of Madness, Love, and the History of the World from the Periodic Table of the ElementsRating: 4 out of 5 stars4/5 (146)
- Monkeys, Myths, and Molecules: Separating Fact from Fiction, and the Science of Everyday LifeFrom EverandMonkeys, Myths, and Molecules: Separating Fact from Fiction, and the Science of Everyday LifeRating: 4 out of 5 stars4/5 (1)
- Essential Oil Chemistry Formulating Essential Oil Blends that Heal - Aldehyde - Ketone - Lactone: Healing with Essential OilFrom EverandEssential Oil Chemistry Formulating Essential Oil Blends that Heal - Aldehyde - Ketone - Lactone: Healing with Essential OilRating: 5 out of 5 stars5/5 (1)
- The Periodic Table of Elements - Post-Transition Metals, Metalloids and Nonmetals | Children's Chemistry BookFrom EverandThe Periodic Table of Elements - Post-Transition Metals, Metalloids and Nonmetals | Children's Chemistry BookNo ratings yet
- The Regenerative Grower's Guide to Garden Amendments: Using Locally Sourced Materials to Make Mineral and Biological Extracts and FermentsFrom EverandThe Regenerative Grower's Guide to Garden Amendments: Using Locally Sourced Materials to Make Mineral and Biological Extracts and FermentsRating: 5 out of 5 stars5/5 (3)
- Handbook of Formulating Dermal Applications: A Definitive Practical GuideFrom EverandHandbook of Formulating Dermal Applications: A Definitive Practical GuideNo ratings yet
- Formulating, Packaging, and Marketing of Natural Cosmetic ProductsFrom EverandFormulating, Packaging, and Marketing of Natural Cosmetic ProductsNo ratings yet
- Science Goes Viral: Captivating Accounts of Science in Everyday LifeFrom EverandScience Goes Viral: Captivating Accounts of Science in Everyday LifeRating: 5 out of 5 stars5/5 (1)
- Bioplastics: A Home Inventors HandbookFrom EverandBioplastics: A Home Inventors HandbookRating: 4 out of 5 stars4/5 (2)
- Phase Equilibria in Chemical EngineeringFrom EverandPhase Equilibria in Chemical EngineeringRating: 4 out of 5 stars4/5 (11)
- AP® Chemistry Crash Course, For the 2020 Exam, Book + Online: Get a Higher Score in Less TimeFrom EverandAP® Chemistry Crash Course, For the 2020 Exam, Book + Online: Get a Higher Score in Less TimeRating: 5 out of 5 stars5/5 (1)
- A Perfect Red: Empire, Espionage, and the Quest for the Color of DesireFrom EverandA Perfect Red: Empire, Espionage, and the Quest for the Color of DesireRating: 4 out of 5 stars4/5 (129)
- Transformer: The Deep Chemistry of Life and DeathFrom EverandTransformer: The Deep Chemistry of Life and DeathRating: 4.5 out of 5 stars4.5/5 (13)