You might also like
- Circadian RhythmsDocument7 pagesCircadian RhythmsSeaton HarnsNo ratings yet
- CST STUDIO SUITE - High Frequency Simulation PDFDocument128 pagesCST STUDIO SUITE - High Frequency Simulation PDFGenik Podunay100% (2)
- DPWH ReviewerDocument597 pagesDPWH Reviewercharles sedigoNo ratings yet
- Gene Regulation of Adult Skeletogenesis in Starfish and Modifications During Gene Network Co OptionDocument13 pagesGene Regulation of Adult Skeletogenesis in Starfish and Modifications During Gene Network Co OptionEzbeidy LemusNo ratings yet
- Vesti - Genetika - Resume Journal Chromosomal MappingDocument3 pagesVesti - Genetika - Resume Journal Chromosomal MappingVesti AtmajaNo ratings yet
- Developmental Evolution: This Side of Paradise: Anthony Graham and Imelda McgonnellDocument3 pagesDevelopmental Evolution: This Side of Paradise: Anthony Graham and Imelda McgonnellAroa ChansNo ratings yet
- Somites Without A Clock 2014 Dias Et AlDocument9 pagesSomites Without A Clock 2014 Dias Et AlCarmen CoelhoNo ratings yet
- A Ubiquitous Spectrolaminar Motif of Local Field Potential Power Across The Primate CortexDocument34 pagesA Ubiquitous Spectrolaminar Motif of Local Field Potential Power Across The Primate CortexTianyi ZhengNo ratings yet
- Varayoud Et Al 2005 ER Alfa Splicing Variants JOEDocument10 pagesVarayoud Et Al 2005 ER Alfa Splicing Variants JOElucasmonje1978No ratings yet
- Vertebrate Ancient-Long Opsin A Green-Sensitive Photoreceptive Molecule Present in Zebrafish Deep Brain and Retinal Horizontal CellsDocument7 pagesVertebrate Ancient-Long Opsin A Green-Sensitive Photoreceptive Molecule Present in Zebrafish Deep Brain and Retinal Horizontal CellsFractalScribd707No ratings yet
- Ceva Cu Sex La SoareciDocument26 pagesCeva Cu Sex La SoareciDragan StefanNo ratings yet
- ERandPR UterusBuffaloDocument11 pagesERandPR UterusBuffalolinda nugrohoNo ratings yet
- Exploring Developmental, Functional, and Evolutionary Aspects of Amphioxus Sensory CellsDocument7 pagesExploring Developmental, Functional, and Evolutionary Aspects of Amphioxus Sensory CellsDefrian MeltaNo ratings yet
- Cortical Activity Early Brain DevelopDocument12 pagesCortical Activity Early Brain DevelopDa CuNo ratings yet
- Evidence For An Expansion-Based Temporal SHH Gradient in Specifying Vertebrate Digit IdentitiesDocument12 pagesEvidence For An Expansion-Based Temporal SHH Gradient in Specifying Vertebrate Digit Identitiesrocambolescas perthNo ratings yet
- Sleep 28 4 395Document16 pagesSleep 28 4 395tserchovNo ratings yet
- Direct Visualization of Nucleogenesis by Precerebellar Neurons: Involvement of Ventricle-Directed, Radial Fibre-Associated MigrationDocument11 pagesDirect Visualization of Nucleogenesis by Precerebellar Neurons: Involvement of Ventricle-Directed, Radial Fibre-Associated Migrationhareb_dNo ratings yet
- Nature PDFDocument23 pagesNature PDFJuliano Rodrigues de CastroNo ratings yet
- Formery 2023 Molecular Evidence of AnteroposteriDocument20 pagesFormery 2023 Molecular Evidence of Anteroposterilei xuNo ratings yet
- A Guide To Assessing Endoplasmic Reticulum HomeostasisDocument16 pagesA Guide To Assessing Endoplasmic Reticulum Homeostasisfernando dominguezNo ratings yet
- Asquith 2004 Tyrosine Phosphorylation ActivatesDocument13 pagesAsquith 2004 Tyrosine Phosphorylation ActivatesGagahNo ratings yet
- Cereb. Cortex-2009-LoTurco-i120-5Document6 pagesCereb. Cortex-2009-LoTurco-i120-5bojun dingNo ratings yet
- Gundlfinger Et Al. - 2010 - Natural Spike Trains Trigger Short - and Long Lasting Dynamics at Hippocampal Mossy Fiber Synapses in RodentsDocument9 pagesGundlfinger Et Al. - 2010 - Natural Spike Trains Trigger Short - and Long Lasting Dynamics at Hippocampal Mossy Fiber Synapses in RodentsGabriel HerreraNo ratings yet
- Segner1994 Article TheDevelopmentOfFunctionalDigeDocument16 pagesSegner1994 Article TheDevelopmentOfFunctionalDigeWicha78No ratings yet
- Efrinas 2Document20 pagesEfrinas 2Jesus MendezNo ratings yet
- ΑιμοποιησηDocument8 pagesΑιμοποιησηIoanna PerakiNo ratings yet
- Delalande1998 PDFDocument3 pagesDelalande1998 PDFNada ZnaidiNo ratings yet
- Ler ThaisDocument6 pagesLer ThaisGustavo FelpeNo ratings yet
- J Bbagrm 2009 02 002Document7 pagesJ Bbagrm 2009 02 002Raymundo Lopez NNo ratings yet
- Developmental Biology: Paula Irles, Saray Ramos, Maria-Dolors PiulachsDocument10 pagesDevelopmental Biology: Paula Irles, Saray Ramos, Maria-Dolors PiulachsMazhar FarNo ratings yet
- Animal Reproduction Science: Characterization of Prostaglandin E Receptors (EP2, EP4) in The Horse OviductDocument7 pagesAnimal Reproduction Science: Characterization of Prostaglandin E Receptors (EP2, EP4) in The Horse OviductDiana RNo ratings yet
- TMP 5 CCDocument13 pagesTMP 5 CCFrontiersNo ratings yet
- EGE Records From Cortical and Deep Brain Structures During Centrifugal and Vibrational Accelerations-hyPDocument7 pagesEGE Records From Cortical and Deep Brain Structures During Centrifugal and Vibrational Accelerations-hyPjulio perezNo ratings yet
- Looking Different: Understanding Diversity in Facial Form: SS 2006 Wiley-Liss, IncDocument9 pagesLooking Different: Understanding Diversity in Facial Form: SS 2006 Wiley-Liss, Incadri steveNo ratings yet
- 2004 PnasDocument6 pages2004 PnasWp SaleNo ratings yet
- A Negative Regulatory Element Required For Light-Dependent Pinopsin Gene ExpressionDocument7 pagesA Negative Regulatory Element Required For Light-Dependent Pinopsin Gene ExpressionFractalScribd707No ratings yet
- Directional Cell Migration Establishes The Axes of Planar Pol - 2004 - DevelopmeDocument12 pagesDirectional Cell Migration Establishes The Axes of Planar Pol - 2004 - Developmehalil sahanNo ratings yet
- Identification and Localization of Estrogen Receptor A-And B-Positive Cells in Adult Male and Female Mouse Intestine at Various Estrogen LevelsDocument7 pagesIdentification and Localization of Estrogen Receptor A-And B-Positive Cells in Adult Male and Female Mouse Intestine at Various Estrogen LevelsChyntia D. RahadiaNo ratings yet
- Art WelDocument10 pagesArt WelWellington Martins Dos SantosNo ratings yet
- Tribolium Segementation Clock 2012 El Sherif Et AlDocument6 pagesTribolium Segementation Clock 2012 El Sherif Et AlCarmen CoelhoNo ratings yet
- 2020 - Porcino - Oviducto Capacitación Espermática PDFDocument19 pages2020 - Porcino - Oviducto Capacitación Espermática PDFMarco Antonio AspronNo ratings yet
- Royer 2020Document12 pagesRoyer 2020nugrahoneyNo ratings yet
- Timing Embryo Segmentation Dynamics and RegulatoryDocument12 pagesTiming Embryo Segmentation Dynamics and RegulatoryHassel Del A PNo ratings yet
- Cortical ControlDocument11 pagesCortical ControlDaviidHernandezNo ratings yet
- JCB 201903033Document20 pagesJCB 201903033Purine ChengNo ratings yet
- 11 Axis FormationDocument61 pages11 Axis Formationapi-3800038100% (1)
- Biol 216 2020 Topic 2 Development - Powerpoint 2.3 Developmental Mechanisms - TaggedDocument45 pagesBiol 216 2020 Topic 2 Development - Powerpoint 2.3 Developmental Mechanisms - TaggedMichael HamoudiNo ratings yet
- Neuroethology of Male Courtship in Drosophila From The GeneDocument14 pagesNeuroethology of Male Courtship in Drosophila From The GeneMert GencNo ratings yet
- U2-Rodriguez-Oliveira Et Al 2022 (Actin Cytoskeleton and Complex Cell Architecture in An Asgard Archaeon)Document26 pagesU2-Rodriguez-Oliveira Et Al 2022 (Actin Cytoskeleton and Complex Cell Architecture in An Asgard Archaeon)LaraKielmasNo ratings yet
- M06 Axisspecificationindrosophila Quad 1Document18 pagesM06 Axisspecificationindrosophila Quad 1Mahendra KatreNo ratings yet
- Evidencia NeurogenesisDocument16 pagesEvidencia NeurogenesisSara Merino EgidoNo ratings yet
- Vpa 25Document11 pagesVpa 25Ada CelisNo ratings yet
- Automatic Detection of Highly Organized Theta Oscillations in The Murine EEGDocument9 pagesAutomatic Detection of Highly Organized Theta Oscillations in The Murine EEGMily Fernanda RamirezNo ratings yet
- Fnhum 14 00063Document9 pagesFnhum 14 00063Fimel LarosaNo ratings yet
- Adachi 2008Document15 pagesAdachi 2008franciscaNo ratings yet
- VPS35 NeurodevelopmentDocument10 pagesVPS35 NeurodevelopmentChunlei WangNo ratings yet
- The Analysis of Brain ActivityDocument4 pagesThe Analysis of Brain ActivityCabinet VeterinarNo ratings yet
- 1 s2.0 S0143400411005339 Main PDFDocument8 pages1 s2.0 S0143400411005339 Main PDFMarco Antonio AspronNo ratings yet
- Rojas 1 s2.0 S2773093X23000168 MainDocument12 pagesRojas 1 s2.0 S2773093X23000168 MainCARLOS PANCHENo ratings yet
- Gravity Sensing in Plant and Animal Cells: NPJ Microgravity December 2021Document11 pagesGravity Sensing in Plant and Animal Cells: NPJ Microgravity December 2021Erickson OngNo ratings yet
- Alsiö Et Al. - 2013 - Ikaros Promotes Early-Born Neuronal Fates in The Cerebral CortexDocument10 pagesAlsiö Et Al. - 2013 - Ikaros Promotes Early-Born Neuronal Fates in The Cerebral CortexXaviBechStruckNo ratings yet
- Characterization of Cognitive Deficits in Rats Overexpressing Human Alpha-Synuclein in The Ventral Tegmental Area and Medial Septum Using Recombinant Adeno-Associated Viral Vectors (2013)Document12 pagesCharacterization of Cognitive Deficits in Rats Overexpressing Human Alpha-Synuclein in The Ventral Tegmental Area and Medial Septum Using Recombinant Adeno-Associated Viral Vectors (2013)Rifqi Fathul ArroisiNo ratings yet
- Reppert and Roode 2018 - HighlightDocument14 pagesReppert and Roode 2018 - HighlightRifqi Fathul ArroisiNo ratings yet
- Comparative Study of The Antioxidant and Enzyme inDocument21 pagesComparative Study of The Antioxidant and Enzyme inRifqi Fathul ArroisiNo ratings yet
- Extraction and Microencapsulation of Tuna Virgin FDocument11 pagesExtraction and Microencapsulation of Tuna Virgin FRifqi Fathul ArroisiNo ratings yet
- s40071 019 0221 1Document13 pagess40071 019 0221 1Rifqi Fathul ArroisiNo ratings yet
- Soil Microbial Resources For Improving Fertilizers Efficiency in An Integrated Plant Nutrient Management SystemDocument25 pagesSoil Microbial Resources For Improving Fertilizers Efficiency in An Integrated Plant Nutrient Management SystemRifqi Fathul ArroisiNo ratings yet
- Microsatellite Loci For The Weaver Ant Oecophylla SmaragdinaDocument4 pagesMicrosatellite Loci For The Weaver Ant Oecophylla SmaragdinaRifqi Fathul ArroisiNo ratings yet
- Molecular Pathogenesis of Shigella SPP.: Controlling Host Cell Signaling, Invasion, and Death by Type III SecretionDocument23 pagesMolecular Pathogenesis of Shigella SPP.: Controlling Host Cell Signaling, Invasion, and Death by Type III SecretionRifqi Fathul ArroisiNo ratings yet
- Genome-Scale Metabolic Modeling Reveals Sars-Cov-2-Induced Metabolic Changes and Antiviral TargetsDocument19 pagesGenome-Scale Metabolic Modeling Reveals Sars-Cov-2-Induced Metabolic Changes and Antiviral TargetsRifqi Fathul ArroisiNo ratings yet
- Animal Behaviour: Alice Bouchard, Klaus ZuberbühlerDocument15 pagesAnimal Behaviour: Alice Bouchard, Klaus ZuberbühlerRifqi Fathul ArroisiNo ratings yet
- Asian Weaver Ants, Oecophylla Smaragdina, and Their Repelling of PollinatorsDocument6 pagesAsian Weaver Ants, Oecophylla Smaragdina, and Their Repelling of PollinatorsRifqi Fathul ArroisiNo ratings yet
- Healthcare 09 01536Document11 pagesHealthcare 09 01536Rifqi Fathul ArroisiNo ratings yet
- Description of A New Species of Microhyla From Bali, Indonesia (Amphibia, Anura)Document13 pagesDescription of A New Species of Microhyla From Bali, Indonesia (Amphibia, Anura)Rifqi Fathul ArroisiNo ratings yet
- Aquatic Ecosystem and Biodiversity A ReviewDocument13 pagesAquatic Ecosystem and Biodiversity A ReviewRifqi Fathul ArroisiNo ratings yet
- Crispr Cas9 Nobel Chemistry November 2020Document34 pagesCrispr Cas9 Nobel Chemistry November 2020Rifqi Fathul ArroisiNo ratings yet
- Shigella Pathogenesis: Molecular and Computational Insights: ReviewDocument23 pagesShigella Pathogenesis: Molecular and Computational Insights: ReviewRifqi Fathul ArroisiNo ratings yet
- Comparative Morphology of Bornean Species of Ansonia (Amphibia Anura Bufonidae) (24pgs)Document24 pagesComparative Morphology of Bornean Species of Ansonia (Amphibia Anura Bufonidae) (24pgs)Rifqi Fathul ArroisiNo ratings yet
- Species Diversity, Elevational Distribution and Reproductive Modes in An Amphibian Community at The Matang Range, Sarawak (Borneo)Document35 pagesSpecies Diversity, Elevational Distribution and Reproductive Modes in An Amphibian Community at The Matang Range, Sarawak (Borneo)Rifqi Fathul ArroisiNo ratings yet
- Ansonia Spinulifer,: Assessment By: IUCN SSC Amphibian Specialist GroupDocument9 pagesAnsonia Spinulifer,: Assessment By: IUCN SSC Amphibian Specialist GroupRifqi Fathul ArroisiNo ratings yet
- Grismer Et Al 2016 Ansonia KhaochangensisDocument26 pagesGrismer Et Al 2016 Ansonia KhaochangensisRifqi Fathul ArroisiNo ratings yet
- Foods 09 00073Document12 pagesFoods 09 00073Rifqi Fathul ArroisiNo ratings yet
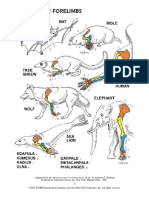
- Adapted From The Human Evolution Coloring Book, 2D Ed., by Adrienne L. Zihlman. Produced by Coloring Concepts Inc. New York: Harpercollins, 2001Document1 pageAdapted From The Human Evolution Coloring Book, 2D Ed., by Adrienne L. Zihlman. Produced by Coloring Concepts Inc. New York: Harpercollins, 2001Rifqi Fathul ArroisiNo ratings yet
- References: Milda Yulia Solihah, 2016Document7 pagesReferences: Milda Yulia Solihah, 2016Rifqi Fathul ArroisiNo ratings yet
- July 2018 1530349036 141Document4 pagesJuly 2018 1530349036 141Rifqi Fathul ArroisiNo ratings yet
- Ijtk 14 (2) 175-182Document8 pagesIjtk 14 (2) 175-182Rifqi Fathul ArroisiNo ratings yet
- Diversity and Floristics of Monocots: - . - Aroids, Lilies, Orchids - .Document66 pagesDiversity and Floristics of Monocots: - . - Aroids, Lilies, Orchids - .Rifqi Fathul ArroisiNo ratings yet
- Conservation of Amphibians and Reptiles in IndonesiaDocument28 pagesConservation of Amphibians and Reptiles in IndonesiaRifqi Fathul ArroisiNo ratings yet
- Clinicopathological Study of Hashimoto Thyroiditis: Pooja Jain, Geetha Vasudevan, Kanthilata PaiDocument5 pagesClinicopathological Study of Hashimoto Thyroiditis: Pooja Jain, Geetha Vasudevan, Kanthilata PaiRifqi Fathul ArroisiNo ratings yet
- Calliandra Paganuccii Leguminosae Mimosoideae A NeDocument5 pagesCalliandra Paganuccii Leguminosae Mimosoideae A NeRifqi Fathul ArroisiNo ratings yet
- Six Types of Species-Area CurvesDocument8 pagesSix Types of Species-Area CurvesRifqi Fathul ArroisiNo ratings yet
- Despiesse de Las Guallas D6H SERIE 3ZF06342Document4 pagesDespiesse de Las Guallas D6H SERIE 3ZF06342David manjarresNo ratings yet
- Guide To Storage Tanks and EquipmentDocument15 pagesGuide To Storage Tanks and EquipmentbadelitamariusNo ratings yet
- 2014 Catbalogan Landslide: September, 17, 2014Document6 pages2014 Catbalogan Landslide: September, 17, 2014Jennifer Gapuz GalletaNo ratings yet
- An Infallible JusticeDocument7 pagesAn Infallible JusticeMani Gopal DasNo ratings yet
- Submittal ReviewDocument22 pagesSubmittal ReviewuddinnadeemNo ratings yet
- Consent For Diagnostic And/or Therapeutic ParacentesisDocument2 pagesConsent For Diagnostic And/or Therapeutic ParacentesisnaveenNo ratings yet
- Gastritis: Department of Gastroenterology General Hospital of Ningxia Medical University Si Cen MDDocument82 pagesGastritis: Department of Gastroenterology General Hospital of Ningxia Medical University Si Cen MDAvi Themessy100% (1)
- Sdo Tle Grade 8 Dressmaking 2nd Q Week 1 8 1Document64 pagesSdo Tle Grade 8 Dressmaking 2nd Q Week 1 8 1Maggie De jesusNo ratings yet
- Handbook (P)Document224 pagesHandbook (P)Joe80% (5)
- Regulatory Framework For Water Dams in QuebecDocument2 pagesRegulatory Framework For Water Dams in QuebecRaveeNo ratings yet
- Notes Ch. 4 - Folk and Popular CultureDocument7 pagesNotes Ch. 4 - Folk and Popular CultureVienna WangNo ratings yet
- Oertel - Extracts From The Jāiminīya-Brāhma A and Upanishad-Brāhma A, Parallel To Passages of TheDocument20 pagesOertel - Extracts From The Jāiminīya-Brāhma A and Upanishad-Brāhma A, Parallel To Passages of Thespongebob2812No ratings yet
- Company ProfileDocument36 pagesCompany ProfileShahzad Nasir SayyedNo ratings yet
- 120 BE5678 CenterDocument2 pages120 BE5678 CenterDipika GuptaNo ratings yet
- Academic Program Required Recommended Academic Program Required RecommendedDocument1 pageAcademic Program Required Recommended Academic Program Required Recommendedonur scribdNo ratings yet
- Chemistry - JEE Main 2024 January - Detailed Analysis by MathonGoDocument3 pagesChemistry - JEE Main 2024 January - Detailed Analysis by MathonGoGulam Naziruddin Makandar (NuTHeaD94)No ratings yet
- Chemical Resistance TableDocument6 pagesChemical Resistance TableEdgarDavidDiazCamposNo ratings yet
- TRL Explanations - 1Document4 pagesTRL Explanations - 1Ana DulceNo ratings yet
- T60n02rg PDFDocument8 pagesT60n02rg PDFsandor9116100% (2)
- A Review of Linear AlgebraDocument19 pagesA Review of Linear AlgebraOsman Abdul-MuminNo ratings yet
- .... Applicant Versus: Cri. Appln. No. 4353/16 & Anr. 1Document11 pages.... Applicant Versus: Cri. Appln. No. 4353/16 & Anr. 1Manashree EngineerNo ratings yet
- Bar Tending TerminologyDocument3 pagesBar Tending TerminologySiska WangNo ratings yet
- Motion of A Simple Pendulum in A FluidDocument16 pagesMotion of A Simple Pendulum in A FluidGokul JeevaNo ratings yet
- Earthing ResistanceDocument4 pagesEarthing ResistanceNeeraj Purohit100% (1)
- The Use of Proton Pump Inhibitors in Treating and Preventing NSAID-induced Mucosal DamageDocument6 pagesThe Use of Proton Pump Inhibitors in Treating and Preventing NSAID-induced Mucosal DamageFriska Rachmanita PrayogoNo ratings yet
- Geology and Mineralization of The Maggie Creek DistrictDocument28 pagesGeology and Mineralization of The Maggie Creek DistrictRockstone Research Ltd.No ratings yet
- Lecture 9: Quarter-Wave-Transformer Matching.: R JZ L Z Z Z JR LDocument13 pagesLecture 9: Quarter-Wave-Transformer Matching.: R JZ L Z Z Z JR LRuth EnormeNo ratings yet
- Big Fat Lies - How The Diet Industry Is Making You Sick, Fat & PoorDocument212 pagesBig Fat Lies - How The Diet Industry Is Making You Sick, Fat & PoorangelobuffaloNo ratings yet