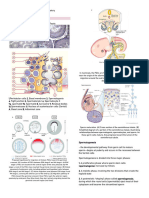
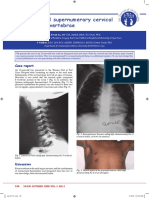
You might also like
- The Anatomy of the Human Peritoneum and Abdominal Cavity: Considered from the Standpoint of Development and Comparative AnatomyFrom EverandThe Anatomy of the Human Peritoneum and Abdominal Cavity: Considered from the Standpoint of Development and Comparative AnatomyNo ratings yet
- 11 Axis FormationDocument61 pages11 Axis Formationapi-3800038100% (1)
- Anatomy of Neuropsychiatry: The New Anatomy of the Basal Forebrain and Its Implications for Neuropsychiatric IllnessFrom EverandAnatomy of Neuropsychiatry: The New Anatomy of the Basal Forebrain and Its Implications for Neuropsychiatric IllnessNo ratings yet
- Science Aax7890 PDFDocument9 pagesScience Aax7890 PDFAndoingNo ratings yet
- Dev151035 PDFDocument19 pagesDev151035 PDFIsnaeni Tri HanifaNo ratings yet
- Effects of Nest Deprivation On Hypothalamic Mesotocin in IncubatingDocument11 pagesEffects of Nest Deprivation On Hypothalamic Mesotocin in Incubatingnatagarn .sNo ratings yet
- Transcribing Pancreas: Perspectives in DiabetesDocument8 pagesTranscribing Pancreas: Perspectives in DiabetesLaurens RingnaldaNo ratings yet
- SHH Signaling Mediated by A Prechordal and Brain Enhancer Controls Forebrain OrganizationDocument7 pagesSHH Signaling Mediated by A Prechordal and Brain Enhancer Controls Forebrain OrganizationPaola Rivera ÁvilaNo ratings yet
- developmental biology notes 3Document26 pagesdevelopmental biology notes 3PrezyNo ratings yet
- Moriyama 2018Document8 pagesMoriyama 2018Gabriel BuonoNo ratings yet
- Advances in Early Kidney DevelopmentDocument12 pagesAdvances in Early Kidney DevelopmentasdNo ratings yet
- 80374f54d421b7b55c189a6a1eeebd628d99Document17 pages80374f54d421b7b55c189a6a1eeebd628d99Ruslan MilevskyNo ratings yet
- 2020 Compr Physiol - Desarrollo de La Glándula PituitariaDocument25 pages2020 Compr Physiol - Desarrollo de La Glándula PituitariaFrancisco PabonNo ratings yet
- Somites Without A Clock 2014 Dias Et AlDocument9 pagesSomites Without A Clock 2014 Dias Et AlCarmen CoelhoNo ratings yet
- The Epididymis Re-Visited: A Personal View: Experience and HistoryDocument6 pagesThe Epididymis Re-Visited: A Personal View: Experience and HistoryAhmad SolihinNo ratings yet
- 1644 FullDocument5 pages1644 FullhermannNo ratings yet
- Peng Nature 2019Document30 pagesPeng Nature 2019Water PowerNo ratings yet
- Bulusu 2017 DevelopmentalCellDocument16 pagesBulusu 2017 DevelopmentalCell22gaefNo ratings yet
- Developmental Dynamics - 2008 - Hu - Effect of Bone Morphogenetic Protein Signaling On Development of The Jaw SkeletonDocument11 pagesDevelopmental Dynamics - 2008 - Hu - Effect of Bone Morphogenetic Protein Signaling On Development of The Jaw Skeletonhennysusanto18No ratings yet
- MMC 5Document27 pagesMMC 5star lightNo ratings yet
- Osificación Endocondral 1Document20 pagesOsificación Endocondral 1Fabian BarretoNo ratings yet
- Segment at I On VertebratesDocument17 pagesSegment at I On VertebratesKeshav JhaNo ratings yet
- (Vincent Et Al, 1997) Ntegration of The Head and Trunk Segmentation Systems Controls Cephalic Furrow Formation in DrosophilaDocument8 pages(Vincent Et Al, 1997) Ntegration of The Head and Trunk Segmentation Systems Controls Cephalic Furrow Formation in DrosophilaエリタノアンソネーNo ratings yet
- Altmann 2001Document36 pagesAltmann 2001zsmm1993No ratings yet
- 73 1189 1 PBDocument2 pages73 1189 1 PBNguyễn Thị Kim TrọngNo ratings yet
- StJohnston 1989Document10 pagesStJohnston 1989Arkha OnNo ratings yet
- Mammalian Cerebral Cortex: Embryonic Development and CytoarchitectureDocument7 pagesMammalian Cerebral Cortex: Embryonic Development and CytoarchitectureDinda Tiara SukmaNo ratings yet
- Actvity # 6 Upper and LOwer ExtremitiesDocument3 pagesActvity # 6 Upper and LOwer ExtremitiesRej CosainNo ratings yet
- Phase Transition in A Cell: C. ElegansDocument3 pagesPhase Transition in A Cell: C. ElegansokafoieahovanNo ratings yet
- Discussion Session 1Document5 pagesDiscussion Session 1Delon van den AkkerNo ratings yet
- Cuello 230329 233416Document14 pagesCuello 230329 233416GaboNo ratings yet
- Obs 03Document33 pagesObs 03Alejo VasquezNo ratings yet
- The Transmembrane Semaphorin Sema I Is Required in Drosophila For Embryonic Motor and CNS Axon GuidanceDocument14 pagesThe Transmembrane Semaphorin Sema I Is Required in Drosophila For Embryonic Motor and CNS Axon GuidancekramkrishnakNo ratings yet
- Recent Advance in Kidney OrganoidsDocument6 pagesRecent Advance in Kidney OrganoidsWilan KrisnaNo ratings yet
- DevbioDocument140 pagesDevbioAaryan GuptaNo ratings yet
- Ezrin Becomes Restricted To Outer Cells Following Asymmetrical Division in The Preimplantation Mouse EmbryoDocument12 pagesEzrin Becomes Restricted To Outer Cells Following Asymmetrical Division in The Preimplantation Mouse EmbryonugrahoneyNo ratings yet
- Consciousness of Body and Mirror Neuron SystemDocument13 pagesConsciousness of Body and Mirror Neuron SystemDare KawatashiNo ratings yet
- 1 s2.0 S1934590911005790 MainDocument8 pages1 s2.0 S1934590911005790 MainReemNo ratings yet
- ZON HematopoyesisDocument16 pagesZON HematopoyesisMariana CabagnaNo ratings yet
- Gene Regulation of Adult Skeletogenesis in Starfish and Modifications During Gene Network Co OptionDocument13 pagesGene Regulation of Adult Skeletogenesis in Starfish and Modifications During Gene Network Co OptionEzbeidy LemusNo ratings yet
- Genome Activation and Architecture in The Early Mammalian EmbryoDocument7 pagesGenome Activation and Architecture in The Early Mammalian EmbryoPetru CernatNo ratings yet
- C ElegansDocument10 pagesC ElegansMmMmGoodNo ratings yet
- Time-Lapse Observation of Stepwise Regression of EDocument10 pagesTime-Lapse Observation of Stepwise Regression of ERifqi Fathul ArroisiNo ratings yet
- tmpB496 TMPDocument17 pagestmpB496 TMPFrontiersNo ratings yet
- Prenatal diagnosis of brainstem anomaliesDocument34 pagesPrenatal diagnosis of brainstem anomaliesVishnu priya kokkulaNo ratings yet
- BIO125ZusammenfassungLF 3Document43 pagesBIO125ZusammenfassungLF 399rnbccdr8No ratings yet
- Early Development in BirdsDocument38 pagesEarly Development in BirdsCM Najito100% (2)
- Chick Embryo DevelopmentDocument5 pagesChick Embryo DevelopmentPaula Manalo-SuliguinNo ratings yet
- Genetics of Axis Specification in DrosophilaDocument3 pagesGenetics of Axis Specification in DrosophilaJohnkevin NocheNo ratings yet
- Bases celulares y TPH (2)Document17 pagesBases celulares y TPH (2)pau_apbNo ratings yet
- Epithelial-Mesenchymal Crosstalk in Wolffian Duct and FetalDocument15 pagesEpithelial-Mesenchymal Crosstalk in Wolffian Duct and FetalEletícia SousaNo ratings yet
- The sense of the physiological condition of the bodyDocument6 pagesThe sense of the physiological condition of the bodyfrompleasureNo ratings yet
- Tsukushi Functions As An Organizer Inducer by Inhibition of - 2004 - DevelopmenDocument12 pagesTsukushi Functions As An Organizer Inducer by Inhibition of - 2004 - Developmenhalil sahanNo ratings yet
- Yothe Growth in Width of The Facial SkeletonDocument6 pagesYothe Growth in Width of The Facial Skeletonalejac1810No ratings yet
- (Harrison Et Al., 2017) Assembly of Embryonic and Extra-Embryonic Stem Cells To Mimic Embryogenesis in VitroDocument24 pages(Harrison Et Al., 2017) Assembly of Embryonic and Extra-Embryonic Stem Cells To Mimic Embryogenesis in VitroJulio dR AltavasNo ratings yet
- GAMETOGENSIS IN HUMANSDocument8 pagesGAMETOGENSIS IN HUMANSKUNAL1221No ratings yet
- Somatic Embryogenesis: An OverviewDocument6 pagesSomatic Embryogenesis: An Overvieweduardo9119No ratings yet
- TMP 6 DAEDocument16 pagesTMP 6 DAEFrontiersNo ratings yet
- 3 FullDocument14 pages3 FullabduolNo ratings yet
- Chapter 18 - Zebrafish Nervous Syste - 2020 - The Zebrafish in Biomedical ResearDocument9 pagesChapter 18 - Zebrafish Nervous Syste - 2020 - The Zebrafish in Biomedical ResearNicolas BaronNo ratings yet
- ANCHETAAADocument79 pagesANCHETAAAGUILAY, KEVIN MARK F.No ratings yet
- Democracy Against Domination PDFDocument257 pagesDemocracy Against Domination PDFvmcosNo ratings yet
- People Vs Sindiong & PastorDocument4 pagesPeople Vs Sindiong & PastorErby Jennifer Sotelo-GesellNo ratings yet
- Quality Improvement Methodologies - PDCA Cycle RAD PDFDocument9 pagesQuality Improvement Methodologies - PDCA Cycle RAD PDFherita suasantaNo ratings yet
- Business Studies Project: Made By: Rahil JainDocument29 pagesBusiness Studies Project: Made By: Rahil JainChirag KothariNo ratings yet
- Facilitating Learner Centered Teaching FinalsDocument3 pagesFacilitating Learner Centered Teaching FinalsPatricia Andrei De GuzmanNo ratings yet
- Group No 5 - Ultratech - Jaypee 20th Sep-1Document23 pagesGroup No 5 - Ultratech - Jaypee 20th Sep-1Snehal100% (1)
- How To Cook Indian: More Than 500 Classic Recipes For The Modern KitchenDocument431 pagesHow To Cook Indian: More Than 500 Classic Recipes For The Modern Kitchenbingoteam100% (1)
- GasGas TXT Racing 2014 Owner's ManualDocument60 pagesGasGas TXT Racing 2014 Owner's Manualstone chouNo ratings yet
- Window Server 2008 Lab ManualDocument51 pagesWindow Server 2008 Lab ManualpadaukzunNo ratings yet
- Slide 2 National Patient SafeDocument34 pagesSlide 2 National Patient Safeapi-3697326No ratings yet
- MTSC QuestionsDocument8 pagesMTSC QuestionsAbhi RamNo ratings yet
- Bitonio V CoaDocument24 pagesBitonio V Coamarjorie requirmeNo ratings yet
- Nilai Murni PKN XII Mipa 3Document8 pagesNilai Murni PKN XII Mipa 3ilmi hamdinNo ratings yet
- FVNL"'" - TT - ..: MycotaDocument8 pagesFVNL"'" - TT - ..: MycotaAmbesh JhaNo ratings yet
- UntitledDocument421 pagesUntitledtunggal KecerNo ratings yet
- Kyle Capodice Ecet Candidate ResumeDocument1 pageKyle Capodice Ecet Candidate Resumeapi-394690479No ratings yet
- Guitar Chord Chart For Drop D TuningDocument4 pagesGuitar Chord Chart For Drop D TuningJnewsletter1No ratings yet
- MSDS TriacetinDocument4 pagesMSDS TriacetinshishirchemNo ratings yet
- Lucid Dreaming Fast TrackDocument3 pagesLucid Dreaming Fast TrackWulan Funblogger50% (2)
- Charles Mwanza's ResumeDocument3 pagesCharles Mwanza's ResumeYash MalemuNo ratings yet
- Sales Confirmation: Alpha Trading S.P.A. Compagnie Tunisienne de NavigationDocument1 pageSales Confirmation: Alpha Trading S.P.A. Compagnie Tunisienne de NavigationimedNo ratings yet
- Partnership Dissolution QuizDocument8 pagesPartnership Dissolution QuizLee SuarezNo ratings yet
- Popular Telugu Novel "Prema DeepikaDocument2 pagesPopular Telugu Novel "Prema Deepikasindhu60% (5)
- Aluminum Extrusions (HFS6 Series) Aluminum Extrusions (HFS6 Series)Document1 pageAluminum Extrusions (HFS6 Series) Aluminum Extrusions (HFS6 Series)Jesus ValenciaNo ratings yet
- Liquichek Specialty Immunoassay Control Levels LTA, 1, 2 and 3Document6 pagesLiquichek Specialty Immunoassay Control Levels LTA, 1, 2 and 3clinicalbiochemistryNo ratings yet
- Revised SCOPE OF WORK and SCCDocument363 pagesRevised SCOPE OF WORK and SCCZia KhanNo ratings yet
- Final test grammar and vocabulary practiceDocument5 pagesFinal test grammar and vocabulary practicexico_euNo ratings yet
- Natural Resources Management IntroDocument14 pagesNatural Resources Management IntroJoaquin David Magana100% (2)
- STELEX PrO (E) FinalDocument4 pagesSTELEX PrO (E) FinalGhita-Mehedintu GheorgheNo ratings yet
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 3.5 out of 5 stars3.5/5 (2)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (3)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (215)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- Eels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishFrom EverandEels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishRating: 4 out of 5 stars4/5 (30)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- Superlative: The Biology of ExtremesFrom EverandSuperlative: The Biology of ExtremesRating: 4.5 out of 5 stars4.5/5 (51)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- The Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineFrom EverandThe Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineRating: 4 out of 5 stars4/5 (17)
- Younger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationFrom EverandYounger for Life: Feel Great and Look Your Best with the New Science of AutojuvenationRating: 4 out of 5 stars4/5 (1)
- Darwin's Dangerous Idea: Evolution and the Meaning of LifeFrom EverandDarwin's Dangerous Idea: Evolution and the Meaning of LifeRating: 4 out of 5 stars4/5 (523)
- Fearfully and Wonderfully: The Marvel of Bearing God's ImageFrom EverandFearfully and Wonderfully: The Marvel of Bearing God's ImageRating: 5 out of 5 stars5/5 (40)
- The Dragons of Eden: Speculations on the Evolution of Human IntelligenceFrom EverandThe Dragons of Eden: Speculations on the Evolution of Human IntelligenceRating: 4 out of 5 stars4/5 (632)