You might also like
- Mathematics 1St First Order Linear Differential Equations 2Nd Second Order Linear Differential Equations Laplace Fourier Bessel MathematicsFrom EverandMathematics 1St First Order Linear Differential Equations 2Nd Second Order Linear Differential Equations Laplace Fourier Bessel MathematicsNo ratings yet
- Lorenz, ChaosDocument342 pagesLorenz, ChaosDaniel Díaz50% (2)
- Delay Differential Equation PDFDocument349 pagesDelay Differential Equation PDFVAIBHAV DHAR DWIVEDINo ratings yet
- (Ben Ayed M., El Mehdi K., Pacella F.) Blow-Up andDocument23 pages(Ben Ayed M., El Mehdi K., Pacella F.) Blow-Up andpaqsoriNo ratings yet
- Abel TheoremxDocument3 pagesAbel TheoremxGerry DundaNo ratings yet
- 10.1007@s11785 019 00924 ZDocument19 pages10.1007@s11785 019 00924 ZKarwan JwamerNo ratings yet
- −1 2 2πiz −b+ √ b −4ac 2a 2 τ 2Document21 pages−1 2 2πiz −b+ √ b −4ac 2a 2 τ 2Sangat BaikNo ratings yet
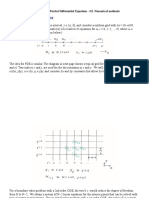
- Numerical Methods For Ode Initial Value ProblemsDocument20 pagesNumerical Methods For Ode Initial Value Problems37 TANNUNo ratings yet
- Eigenvalues To DiagonalizationDocument21 pagesEigenvalues To DiagonalizationMajety S LskshmiNo ratings yet
- Diagonal Dominance and Harmless Off-Diagonal DelaysDocument8 pagesDiagonal Dominance and Harmless Off-Diagonal DelaysMark Edward GonzalesNo ratings yet
- On Complex Extrapolated Successive Overrelaxation (Esor) : Some Theoretical ResultsDocument16 pagesOn Complex Extrapolated Successive Overrelaxation (Esor) : Some Theoretical Resultslight crow songNo ratings yet
- Tmna 2016 043Document21 pagesTmna 2016 043Steffanio MorenoNo ratings yet
- Module2 ADDE (MAT2002)Document27 pagesModule2 ADDE (MAT2002)GURU PRASATHNo ratings yet
- Ma1002: (Ordinary Differential Equations) : Part-I: First Oder OdesDocument31 pagesMa1002: (Ordinary Differential Equations) : Part-I: First Oder OdesVishal KumarNo ratings yet
- Spiral Singularities of A SemiflowDocument14 pagesSpiral Singularities of A SemiflowKamil DunstNo ratings yet
- Sturm-Liouville Problems: Generalized Fourier Series: X X (0) 0, X (L) 0Document13 pagesSturm-Liouville Problems: Generalized Fourier Series: X X (0) 0, X (L) 0Sebastian ReguanNo ratings yet
- On The Accuracy of Grid Approximations To Nonsmooth Solutions of A Singularly Perturbed Reaction-Diffusion Equation in The SquareDocument13 pagesOn The Accuracy of Grid Approximations To Nonsmooth Solutions of A Singularly Perturbed Reaction-Diffusion Equation in The Squarekaushik9282No ratings yet
- CEE 471, Fall 2019: HW9 SolutionsDocument12 pagesCEE 471, Fall 2019: HW9 SolutionsJonathanNo ratings yet
- Reduced Words and Plane Partitions: 1. Main ResultDocument9 pagesReduced Words and Plane Partitions: 1. Main Resultnia rili putri irawanNo ratings yet
- 14-Method of Variation of Parameter-05!09!2019 (05-Sep-2019) Material I 05-Sep-2019 Module 3.4 VariatDocument2 pages14-Method of Variation of Parameter-05!09!2019 (05-Sep-2019) Material I 05-Sep-2019 Module 3.4 VariatParth VijayNo ratings yet
- Estimation of Parametric Functions in Downton'sDocument17 pagesEstimation of Parametric Functions in Downton'sRacha A AliNo ratings yet
- Generating Random VariablesDocument7 pagesGenerating Random VariablesTsvetanNo ratings yet
- Lecture 5Document15 pagesLecture 52220678No ratings yet
- HW 13 ADocument8 pagesHW 13 AAjay Varma100% (5)
- Lie Groups, Lie Algebras, and Their RepresentationsDocument85 pagesLie Groups, Lie Algebras, and Their RepresentationssharlineNo ratings yet
- Introduction To Partial Differential Equation - III. Numerical MethodsDocument18 pagesIntroduction To Partial Differential Equation - III. Numerical Methodssoundarya raghuwanshiNo ratings yet
- CharacteristicDocument13 pagesCharacteristicvictor bravoNo ratings yet
- Disproof of A Conjecture by Rademacher On Partial Fractions: X (1, - . - , N) ), X C EDocument14 pagesDisproof of A Conjecture by Rademacher On Partial Fractions: X (1, - . - , N) ), X C EshyamNo ratings yet
- MATH 304 Linear Algebra Matrix Transformations (Continued) - Matrix of A Linear TransformationDocument26 pagesMATH 304 Linear Algebra Matrix Transformations (Continued) - Matrix of A Linear TransformationUp ToyouNo ratings yet
- Integral Equations For Special Functions of Heun CDocument12 pagesIntegral Equations For Special Functions of Heun Cir.noselidzeNo ratings yet
- Department of Mathematics University of Petroleum & Energy Studies Assignment-IDocument2 pagesDepartment of Mathematics University of Petroleum & Energy Studies Assignment-Iananthu.uNo ratings yet
- Direct Method SlidesDocument44 pagesDirect Method SlideszazarNo ratings yet
- Lesson2-2 (2) - 2 PDFDocument11 pagesLesson2-2 (2) - 2 PDFHugo NavaNo ratings yet
- 221B Lecture Notes: Notes On Spherical Bessel Functions 1 DefinitionsDocument5 pages221B Lecture Notes: Notes On Spherical Bessel Functions 1 DefinitionsKhem UpathambhakulNo ratings yet
- BellewDocument16 pagesBellewCander CyNo ratings yet
- The Legendre Equation and Its Self-AdjointDocument33 pagesThe Legendre Equation and Its Self-AdjointmohammadH7No ratings yet
- Ps 9 SolDocument2 pagesPs 9 SolAyman FansaNo ratings yet
- New Interesting Euler Sums: Ournal of Lassical Nalysis Volume 15, Number 1 (2019), 9-22Document14 pagesNew Interesting Euler Sums: Ournal of Lassical Nalysis Volume 15, Number 1 (2019), 9-22Papy MaikaNo ratings yet
- Some Non-Local Problems For The Parabolic-Hyperbolic Type Equation With Non-Characteristic Line of Changing TypeDocument11 pagesSome Non-Local Problems For The Parabolic-Hyperbolic Type Equation With Non-Characteristic Line of Changing TypeErkinjonNo ratings yet
- Dyadic Wavelets and Refinable Functions On A Half-LineDocument30 pagesDyadic Wavelets and Refinable Functions On A Half-LineukoszapavlinjeNo ratings yet
- HW 11Document3 pagesHW 11snow92686No ratings yet
- Statistics and Finance: An: David RuppertDocument46 pagesStatistics and Finance: An: David Ruppertmastanoob50% (2)
- Castaneda and Berres (2023) - Bifurcation of Solutions Through A Contact Manifold in Bidisperse ModelsDocument4 pagesCastaneda and Berres (2023) - Bifurcation of Solutions Through A Contact Manifold in Bidisperse ModelsPablo CastañedaNo ratings yet
- Math 121A: Solutions To Final Exam: AnswerDocument12 pagesMath 121A: Solutions To Final Exam: AnswercfisicasterNo ratings yet
- Lay Slavyanov 1999Document16 pagesLay Slavyanov 1999ir.noselidzeNo ratings yet
- EM703p Higher Order Linear ODEsDocument10 pagesEM703p Higher Order Linear ODEsDHT845No ratings yet
- Higher Order ODEDocument30 pagesHigher Order ODESumukh KiniNo ratings yet
- Initial-Boundary Value Problems For The Wave Equation: Electronic Journal of Differential Equations February 2014Document7 pagesInitial-Boundary Value Problems For The Wave Equation: Electronic Journal of Differential Equations February 2014Batman BatmanNo ratings yet
- The K Ahler Geometry of Toric Manifolds: E-Mail Address: Apostolov - Vestislav@uqam - CaDocument61 pagesThe K Ahler Geometry of Toric Manifolds: E-Mail Address: Apostolov - Vestislav@uqam - CaIvan Coronel MarthensNo ratings yet
- MATH 219: Spring 2021-22Document6 pagesMATH 219: Spring 2021-22HesapNo ratings yet
- A Comment On Methods To Deal With Divergent Series and IntegralsDocument14 pagesA Comment On Methods To Deal With Divergent Series and Integralseljose1234100% (2)
- Jordan SuperalgebraDocument13 pagesJordan SuperalgebraGulnoza Bo'stonovaNo ratings yet
- Lect2 AgmDocument63 pagesLect2 AgmShu Shujaat LinNo ratings yet
- Problems and Solutions in Differential Geometry and ApplicationsDocument125 pagesProblems and Solutions in Differential Geometry and ApplicationsseimomNo ratings yet
- On Imbedding Theorems For Weighted Anisotropic Sobolev SpacesDocument13 pagesOn Imbedding Theorems For Weighted Anisotropic Sobolev SpacesLuis Alberto FuentesNo ratings yet
- Miersemann - Elliptic Partial Differential Equations of Second OrderDocument87 pagesMiersemann - Elliptic Partial Differential Equations of Second OrderCristian ValdezNo ratings yet
- Fundamental Solution Set in Case of Repeated Eigen ValueDocument5 pagesFundamental Solution Set in Case of Repeated Eigen ValueJimmy WernerNo ratings yet
- LCT 4Document30 pagesLCT 4Sandipan SahaNo ratings yet
- Tables of the Function w (z)- e-z2 ? ex2 dx: Mathematical Tables Series, Vol. 27From EverandTables of the Function w (z)- e-z2 ? ex2 dx: Mathematical Tables Series, Vol. 27No ratings yet
- Tables of Generalized Airy Functions for the Asymptotic Solution of the Differential Equation: Mathematical Tables SeriesFrom EverandTables of Generalized Airy Functions for the Asymptotic Solution of the Differential Equation: Mathematical Tables SeriesNo ratings yet
- Evaluation of MATCONT Bifurcation - W Jason PicardoDocument10 pagesEvaluation of MATCONT Bifurcation - W Jason PicardoFatima Muhammad SaleemNo ratings yet
- Mth744u Exam 2013Document3 pagesMth744u Exam 2013Panos SiatosNo ratings yet
- Vortex DynamicsDocument15 pagesVortex DynamicsAditya KumarNo ratings yet
- Matlab-Based Voltage Stability AnalysisDocument5 pagesMatlab-Based Voltage Stability AnalysisSharsu GadgetNo ratings yet
- ACMTOMS HomDocument35 pagesACMTOMS HomPetar ĆirkovićNo ratings yet
- Prediction of Voltage Stability in Power System by Using CPF MethodDocument6 pagesPrediction of Voltage Stability in Power System by Using CPF MethodijsretNo ratings yet
- AnswerExam 2010 02Document15 pagesAnswerExam 2010 02Snigdha Thakur RaiNo ratings yet
- Bifurcation Analysis of The Modified Lotka-Volterra Prey-Predator ModelDocument44 pagesBifurcation Analysis of The Modified Lotka-Volterra Prey-Predator ModelMiliyon TilahunNo ratings yet
- Transcritical BifurcationDocument16 pagesTranscritical BifurcationDave MillingtonNo ratings yet
- Investigation of Wind Farm On Power System Voltage Stability Based On Bifurcation TheoryDocument4 pagesInvestigation of Wind Farm On Power System Voltage Stability Based On Bifurcation TheoryArmando MaloneNo ratings yet
- The Broken Window FallacyDocument9 pagesThe Broken Window Fallacybjuti folkNo ratings yet
- Fracmemristor Chaotic Oscillator With MultistableDocument15 pagesFracmemristor Chaotic Oscillator With MultistablepunithNo ratings yet
- HW 1Document5 pagesHW 1tt_aljobory3911No ratings yet
- Bifurcation Theory ThesisDocument67 pagesBifurcation Theory ThesisJohn StarrettNo ratings yet
- Homework Set 3 Some Solutions: Course Web Site: HTTP://WWW - Math.sfu - Ca/ Ralfw/math467Document8 pagesHomework Set 3 Some Solutions: Course Web Site: HTTP://WWW - Math.sfu - Ca/ Ralfw/math467Annas FauzyNo ratings yet
- Bifurcation TheoryDocument13 pagesBifurcation Theoryapi-491799610No ratings yet
- Nikolova 2020Document9 pagesNikolova 2020cesar abraham torrico chavezNo ratings yet
- Taylor SeriesDocument9 pagesTaylor SeriesAbdul Wasiq KhanNo ratings yet
- Patidar Nonstandard Finite Difference Methods PDFDocument34 pagesPatidar Nonstandard Finite Difference Methods PDFjjj_ddd_pierreNo ratings yet
- Border-Collision Bifurcations in The Buck ConverterDocument10 pagesBorder-Collision Bifurcations in The Buck ConverterNéstor BernalNo ratings yet
- Shimmy in Aircraft Landing Gear: Etienne CoetzeeDocument11 pagesShimmy in Aircraft Landing Gear: Etienne CoetzeerodefyNo ratings yet
- Final Power ComponentDocument22 pagesFinal Power ComponentAbdullah AzizNo ratings yet
- Bifurcation Theory and Its Application T PDFDocument5 pagesBifurcation Theory and Its Application T PDFmarrhewNo ratings yet
- Che614 Introduction To Hydrodynamic Stability Assignment 2 Due Date: 18 August 2014 Elementary Theory of BifurcationsDocument1 pageChe614 Introduction To Hydrodynamic Stability Assignment 2 Due Date: 18 August 2014 Elementary Theory of BifurcationsAnnas FauzyNo ratings yet
- Chaos in Lotka Volterra ModelsDocument14 pagesChaos in Lotka Volterra Modelsjjj_ddd_pierreNo ratings yet
- Gregory S. Ezra - Classical-Quantum Correspondence and The Analysis of Highly-Excited States: Periodic Orbits, Rational Tori and BeyondDocument49 pagesGregory S. Ezra - Classical-Quantum Correspondence and The Analysis of Highly-Excited States: Periodic Orbits, Rational Tori and BeyondOmasazzNo ratings yet
- Tutorial PDFDocument10 pagesTutorial PDFmaheshnagarkarNo ratings yet
- Differential Equations (AIMS 2015)Document103 pagesDifferential Equations (AIMS 2015)Robin Red Msiska0% (1)