You might also like
- Instant Download Ebook PDF Biopsychology 9th Edition by John P J Pinel PDF ScribdDocument41 pagesInstant Download Ebook PDF Biopsychology 9th Edition by John P J Pinel PDF Scribdfranklin.mayer855100% (38)
- Osmosis High-Yield Physiology - Medicalstudyzone - Com (Sürüklenen)Document87 pagesOsmosis High-Yield Physiology - Medicalstudyzone - Com (Sürüklenen)Dilay MercimekNo ratings yet
- Neurological MCQDocument4 pagesNeurological MCQAnonymous hF5zAdvwCC100% (1)
- Spinal Cord InjuryDocument37 pagesSpinal Cord InjuryMaryanneNo ratings yet
- Fear and Anxiety - Animal Models and Human Cognitive Psychophysiology PDFDocument23 pagesFear and Anxiety - Animal Models and Human Cognitive Psychophysiology PDFNST MtzNo ratings yet
- Anxiety Animals ModelsDocument9 pagesAnxiety Animals ModelsNST MtzNo ratings yet
- SCIENCE 10-DLP1-Nervous-SystemDocument3 pagesSCIENCE 10-DLP1-Nervous-SystemSharlene Cecil PagoboNo ratings yet
- Abnormalities of Neuromuscular Transmission in Patients With Miller-Fisher SyndromeDocument3 pagesAbnormalities of Neuromuscular Transmission in Patients With Miller-Fisher SyndromearaliNo ratings yet
- Etm 20 05 09218Document7 pagesEtm 20 05 09218José A. RodríguezNo ratings yet
- Neuronal Activity and Outcomes From Thalamic Surgery For Spinocerebellar AtaxiaDocument12 pagesNeuronal Activity and Outcomes From Thalamic Surgery For Spinocerebellar AtaxiaNiken HapsariNo ratings yet
- Maeshima y Osawa - 2007 - Stroke Rehabilitation in A Patient With CerebellarDocument8 pagesMaeshima y Osawa - 2007 - Stroke Rehabilitation in A Patient With CerebellarJEREMY RANo ratings yet
- França (2018) Brain StimulationDocument12 pagesFrança (2018) Brain StimulationAna paula CamargoNo ratings yet
- ReviewDocument13 pagesReviewSnezana MihajlovicNo ratings yet
- Spect PDFDocument14 pagesSpect PDFhalamadrid77No ratings yet
- 388 FullDocument7 pages388 Fullchartreuse avonleaNo ratings yet
- Episodic Astasia-Abasia Associated With Hyper-Perfusion in The Subthalamic Region and Dorsal BrainstemDocument3 pagesEpisodic Astasia-Abasia Associated With Hyper-Perfusion in The Subthalamic Region and Dorsal BrainstemAnonymous vnv6QFNo ratings yet
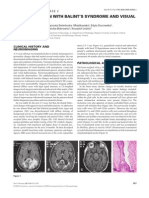
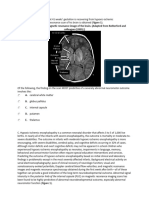
- 31-Year-Old Man With Balint'S Syndrome and Visual Problems: Clinical History and NeuroimagingDocument4 pages31-Year-Old Man With Balint'S Syndrome and Visual Problems: Clinical History and NeuroimagingwillygopeNo ratings yet
- Basal Ganglia LesionsDocument4 pagesBasal Ganglia LesionsPeter Francis RaguindinNo ratings yet
- Bimbingan BsDocument42 pagesBimbingan BsMohamad NasrullohNo ratings yet
- Tugas Bedah Saraf: Pembimbing: Dr. Arinta Puspita Wati, Sps Oleh: Astri Angelina SitaniapessyDocument42 pagesTugas Bedah Saraf: Pembimbing: Dr. Arinta Puspita Wati, Sps Oleh: Astri Angelina SitaniapessyMohamad NasrullohNo ratings yet
- Desordenes Musculares GatosDocument13 pagesDesordenes Musculares GatosDulce BorbollaNo ratings yet
- Carrasco Moro R Beyond Uncal Herniation An UpdatedDocument22 pagesCarrasco Moro R Beyond Uncal Herniation An UpdatedRaul de Sousa RamosNo ratings yet
- MuscularDocument5 pagesMuscularAdFirNo ratings yet
- Stroke 2013 Hinman 182 9Document8 pagesStroke 2013 Hinman 182 9SalwiyadiJarwisJaminNo ratings yet
- Posterior Reversible Encephalopathy Syndrome After Lumbar PunctureDocument2 pagesPosterior Reversible Encephalopathy Syndrome After Lumbar PunctureMedtext PublicationsNo ratings yet
- tmp783C TMPDocument1 pagetmp783C TMPFrontiersNo ratings yet
- Motor Neuron Degeneration Correlates With Respiratory Dysfunction in SCA1Document7 pagesMotor Neuron Degeneration Correlates With Respiratory Dysfunction in SCA1Arie AnggaNo ratings yet
- Awards Works: Expanded AbstractDocument2 pagesAwards Works: Expanded AbstractJelvis BofNo ratings yet
- JNEUROSCIENCEDocument40 pagesJNEUROSCIENCERodrigo Aguirre BáezNo ratings yet
- Brain Pathology - January 1994 - Johansson - Functional Recovery After Brain Infarction Plasticity and NeuralDocument11 pagesBrain Pathology - January 1994 - Johansson - Functional Recovery After Brain Infarction Plasticity and NeuralMauro CracchioloNo ratings yet
- Neurology 2013 Agayeva E119Document2 pagesNeurology 2013 Agayeva E119Luther ThengNo ratings yet
- Chat 5HT Brain Stem Mouse AnatomyDocument35 pagesChat 5HT Brain Stem Mouse AnatomyRam KNo ratings yet
- Anatomia y Fisiologia de GB MDSDocument9 pagesAnatomia y Fisiologia de GB MDSMauricioNo ratings yet
- NeoPREP 2011Document342 pagesNeoPREP 2011johnbarrow641No ratings yet
- Neuroasia 2017 22 (3) 185Document7 pagesNeuroasia 2017 22 (3) 185Ray ParikesitNo ratings yet
- Neural Correlates of Cerebral Plasticity After Brain InfarctionDocument5 pagesNeural Correlates of Cerebral Plasticity After Brain Infarctionarif 2006No ratings yet
- Workshop E - Bundle 2019 - Electro-274-295Document22 pagesWorkshop E - Bundle 2019 - Electro-274-295adaptacion neonatal100% (1)
- C R B S: Urrent Eview in Asic CienceDocument5 pagesC R B S: Urrent Eview in Asic CienceNurul FajriNo ratings yet
- E194 FullDocument3 pagesE194 FullAlex BabyNo ratings yet
- SdarticleDocument2 pagesSdarticlealysandrNo ratings yet
- Cortical Laminar NecrosisDocument7 pagesCortical Laminar Necrosisbalajinagarajanas2001No ratings yet
- FullDocument8 pagesFullrocambolescas perthNo ratings yet
- Dysmetria of Thought Clinical Consequences of CerDocument12 pagesDysmetria of Thought Clinical Consequences of Cervikhen0712No ratings yet
- Krabbe's ImagingDocument4 pagesKrabbe's Imagingshruti kumarNo ratings yet
- NatureDocument10 pagesNatureEduardo Blanco CalvoNo ratings yet
- J Clinph 2013 03 031Document7 pagesJ Clinph 2013 03 031Andres Rojas JerezNo ratings yet
- Plexiform Neurofibroma of The Cauda Equina Presenting As Peroneal Muscular Atrophy (Muscle & Nerve, Vol. 19, Issue 2) (1996)Document4 pagesPlexiform Neurofibroma of The Cauda Equina Presenting As Peroneal Muscular Atrophy (Muscle & Nerve, Vol. 19, Issue 2) (1996)antonioopNo ratings yet
- Quartarone 2013Document10 pagesQuartarone 2013GsNo ratings yet
- Role of MLC901 in Increasing Neurogenesis in Rats With Traumatic Brain InjuryDocument5 pagesRole of MLC901 in Increasing Neurogenesis in Rats With Traumatic Brain InjuryInggitaDarmawanNo ratings yet
- Encefalitis LimbicaDocument11 pagesEncefalitis LimbicaRandy UlloaNo ratings yet
- The Shrinking Brain: Cerebral Atrophy Following Traumatic Brain InjuryDocument19 pagesThe Shrinking Brain: Cerebral Atrophy Following Traumatic Brain InjuryidkNo ratings yet
- Neuromonitoring SyllabusDocument9 pagesNeuromonitoring SyllabuspablocotanNo ratings yet
- A 10-Year Old Girl With Neck Pain: Clinical HistoryDocument4 pagesA 10-Year Old Girl With Neck Pain: Clinical HistorywillygopeNo ratings yet
- Double Crush SyndromeDocument6 pagesDouble Crush SyndromemesrickNo ratings yet
- The Value of Electromyography: NeurologyDocument2 pagesThe Value of Electromyography: NeurologyAnonymous ovTI4eNo ratings yet
- Neuro Qu Mic Adela EpilepsiaDocument10 pagesNeuro Qu Mic Adela EpilepsiaKatiuska Yaritza Renteria ValdiviesoNo ratings yet
- Traumatic Medial Temporal Lobe Epilepsy Associated With A Subcortical Lesion in The Internal Capsule - A Case ReportDocument0 pagesTraumatic Medial Temporal Lobe Epilepsy Associated With A Subcortical Lesion in The Internal Capsule - A Case ReportAyu Ersya WindiraNo ratings yet
- Mechanisms of B-WPS OfficeDocument22 pagesMechanisms of B-WPS OfficeDaylan Lindo MontefalcoNo ratings yet
- (10920684 - Neurosurgical Focus) Factors Affecting Neurological Outcome in Traumatic Conus Medullaris and Cauda Equina InjuriesDocument9 pages(10920684 - Neurosurgical Focus) Factors Affecting Neurological Outcome in Traumatic Conus Medullaris and Cauda Equina InjuriesEddy SaadNo ratings yet
- Neuroimagen en La Esclerosis Lateral Amiotrófica Usos Actuales y EmergentesDocument34 pagesNeuroimagen en La Esclerosis Lateral Amiotrófica Usos Actuales y EmergentesNatalia VelezNo ratings yet
- 0004 282X Anp 72 06 451Document6 pages0004 282X Anp 72 06 451Okhuarobo SamuelNo ratings yet
- Hemimegalencephaly: A Study of Abnormalities Occurring Outside The Involved HemisphereDocument5 pagesHemimegalencephaly: A Study of Abnormalities Occurring Outside The Involved Hemisphereapi-148100258No ratings yet
- Neuroimage: ClinicalDocument7 pagesNeuroimage: ClinicalRendra Syani Ulya FitriNo ratings yet
- Acutecentralnervous Systemtraumainthefield: Krista EstellDocument14 pagesAcutecentralnervous Systemtraumainthefield: Krista EstellJuan Pardo CarrascoNo ratings yet
- Journal of Chemical NeuroanatomyDocument1 pageJournal of Chemical NeuroanatomyNST MtzNo ratings yet
- Whatever Happened To Little AlbertDocument51 pagesWhatever Happened To Little AlbertNST MtzNo ratings yet
- Mouse Monoclonal Antibody Synaptophysin: Novocastra LiquidDocument2 pagesMouse Monoclonal Antibody Synaptophysin: Novocastra LiquidNST MtzNo ratings yet
- Whatever Happened To Little Albert PDFDocument10 pagesWhatever Happened To Little Albert PDFNST MtzNo ratings yet
- Role of Adrenoceptor Subtypes in Memory ConsolidationDocument47 pagesRole of Adrenoceptor Subtypes in Memory ConsolidationNST MtzNo ratings yet
- Cognitive Assesment of OCD PDFDocument15 pagesCognitive Assesment of OCD PDFNST MtzNo ratings yet
- Neurosciencie For KidsDocument10 pagesNeurosciencie For KidsAna Maria RodriguezNo ratings yet
- DraganskiDocument5 pagesDraganskikelly doNo ratings yet
- Nervous System (Parts & Functions)Document30 pagesNervous System (Parts & Functions)Sharlene Cecil PagoboNo ratings yet
- 1st SemDocument158 pages1st SemSana chaudharyNo ratings yet
- PSYC Consciousness NotesDocument3 pagesPSYC Consciousness NotesPhiwe MajokaneNo ratings yet
- Chapter 19 Introduction To Nerves and The Nervous SystemDocument27 pagesChapter 19 Introduction To Nerves and The Nervous SystemFugado Mica illaNo ratings yet
- Neuroelectrophysiology Paper 2Document6 pagesNeuroelectrophysiology Paper 2Harveen BawaNo ratings yet
- Tercer Informe de FisiologiaDocument6 pagesTercer Informe de FisiologiaYanet Onofre PilcoNo ratings yet
- Blakemore and CooperDocument2 pagesBlakemore and Coopermeenaxi.aravinthanNo ratings yet
- Quiz#4Document3 pagesQuiz#4jhacetin775No ratings yet
- Principles of Animal Physiology 3Rd Edition Moyes Test Bank Full Chapter PDFDocument35 pagesPrinciples of Animal Physiology 3Rd Edition Moyes Test Bank Full Chapter PDFAndrewRobinsonixez100% (7)
- Mah 2020Document2 pagesMah 2020Andana TrisaviNo ratings yet
- Chapter 3 - Biology & NeuroscienceDocument64 pagesChapter 3 - Biology & NeuroscienceakshitaNo ratings yet
- SLMCCM Self Study Guide 1 BerDocument3 pagesSLMCCM Self Study Guide 1 BerPaolo PanlaquiNo ratings yet
- HHG4M Artefact 311 Challenge1Document3 pagesHHG4M Artefact 311 Challenge1Maddison VangNo ratings yet
- The Nervous System 1ae60 62e99ab3Document1 pageThe Nervous System 1ae60 62e99ab3shamshadNo ratings yet
- Martella Et Al.Document7 pagesMartella Et Al.Daniela SalazarNo ratings yet
- Q2 Week 4 Day 2Document4 pagesQ2 Week 4 Day 2chona redillasNo ratings yet
- L 1 Unit 1 Medicinal Chemistry LL LATEST LEARN PHARMACYDocument5 pagesL 1 Unit 1 Medicinal Chemistry LL LATEST LEARN PHARMACYYASH DESHMUKH 66No ratings yet
- Vanders Human Physiology The Mechanisms of Body Function 14Th Edition Widmaier Solutions Manual Full Chapter PDFDocument44 pagesVanders Human Physiology The Mechanisms of Body Function 14Th Edition Widmaier Solutions Manual Full Chapter PDFgadsmanoutfitqcs100% (5)
- Cognitive Psychology and Its Implications 8th Edition Anderson Test BankDocument21 pagesCognitive Psychology and Its Implications 8th Edition Anderson Test Bankalexispatrickespgjiyntd100% (32)
- Unit 3Document62 pagesUnit 3Rishika ShrivastavaNo ratings yet
- Modern Essentials - 12th EditionDocument12 pagesModern Essentials - 12th Editionaura.ghimisNo ratings yet
- Cursus Introductie-2324Document81 pagesCursus Introductie-2324alu0101474519No ratings yet
- Autonomic CH 15Document34 pagesAutonomic CH 15sawalekatumbo1No ratings yet
- Local Anesthesia For The Dental Hygienist 1st Edition Logothetis Test BankDocument11 pagesLocal Anesthesia For The Dental Hygienist 1st Edition Logothetis Test Bankcatherineriosidjt100% (24)
- Speaker Notes PsychDocument2 pagesSpeaker Notes PsychkeiramaryhillNo ratings yet