You might also like
- Cellular Dynamics of the Neuron: Symposia of the International Society for Cell Biology, Vol. 8From EverandCellular Dynamics of the Neuron: Symposia of the International Society for Cell Biology, Vol. 8No ratings yet
- Arroyo_J_Neurosci_1999Document13 pagesArroyo_J_Neurosci_1999Constanza FuentesNo ratings yet
- Carr Central ProjectionsDocument13 pagesCarr Central ProjectionsMengda ZhangNo ratings yet
- Effect of Nerve Crush On Perikaryal Number and Volume of Neurons in Adult Rat Dorsal Root GanglionDocument7 pagesEffect of Nerve Crush On Perikaryal Number and Volume of Neurons in Adult Rat Dorsal Root Ganglionrnnr2159No ratings yet
- Edwards - Neck Muscle Afferents Influence Oromotor and Cardiorespiratory CircuitsDocument16 pagesEdwards - Neck Muscle Afferents Influence Oromotor and Cardiorespiratory CircuitssbachorickNo ratings yet
- Seminario 2Document20 pagesSeminario 2LeonardoNo ratings yet
- High-Resolution Measurement of Electrically-Evoked Vagus Nerve Activity in The Anesthetized DogDocument10 pagesHigh-Resolution Measurement of Electrically-Evoked Vagus Nerve Activity in The Anesthetized Dogsuper_montyNo ratings yet
- Fncel 08 00148Document18 pagesFncel 08 00148FabricioNo ratings yet
- Jphysiol00385 0110Document9 pagesJphysiol00385 0110MichaelNo ratings yet
- Braistem ReflectsDocument9 pagesBraistem ReflectsPedro Waskar Aramayo TorricoNo ratings yet
- Nokura 1997Document7 pagesNokura 1997Luis CsrNo ratings yet
- Genetic Comparison of Seizure Control by Norepinephrine and Neuropeptide YDocument6 pagesGenetic Comparison of Seizure Control by Norepinephrine and Neuropeptide YFrontiersNo ratings yet
- Natural Waking and Sleep State A View From Inside Neocortical NeuronsDocument17 pagesNatural Waking and Sleep State A View From Inside Neocortical NeuronsLeslie AcevesNo ratings yet
- Mironov 2000Document7 pagesMironov 20000hitk0No ratings yet
- Schizophrenic Patients. Abnormal Auditory Brain-Stem Responses in HallucinatingDocument7 pagesSchizophrenic Patients. Abnormal Auditory Brain-Stem Responses in HallucinatingHadzrie HamdanNo ratings yet
- Ernfors P, Kucera J, Lee KF, Et Al. (1995)Document9 pagesErnfors P, Kucera J, Lee KF, Et Al. (1995)Rusda AnandaNo ratings yet
- Experimental Physiology - 2004 - O Leary - Discharge Patterns of Preganglionic Neurones With Axons in A Cardiac VagalDocument13 pagesExperimental Physiology - 2004 - O Leary - Discharge Patterns of Preganglionic Neurones With Axons in A Cardiac VagalAndré DiasNo ratings yet
- Ouantitative Assessment of Tactile Allodynia in The Rat PawDocument9 pagesOuantitative Assessment of Tactile Allodynia in The Rat PawAskarNo ratings yet
- 3 Muscle Sympatethic InnervationDocument7 pages3 Muscle Sympatethic InnervationGeraldi ContrerasNo ratings yet
- Criteria For Diagnosis of Pure Neural Leprosy: Original CommunicationDocument4 pagesCriteria For Diagnosis of Pure Neural Leprosy: Original CommunicationFAZRI MONo ratings yet
- Delorme 1997Document8 pagesDelorme 1997groot marvelNo ratings yet
- Hefti - 1988Document13 pagesHefti - 1988Majd HusseinNo ratings yet
- Reference-Free Quantification of EEG Spectra: Combining Current Source Density (CSD) and Frequency Principal Components Analysis (fPCA)Document21 pagesReference-Free Quantification of EEG Spectra: Combining Current Source Density (CSD) and Frequency Principal Components Analysis (fPCA)Raynald SumampouwNo ratings yet
- Kitao 1996Document9 pagesKitao 1996M4shroomNo ratings yet
- Jneurosci 0073-14 2014Document15 pagesJneurosci 0073-14 2014mahfuz emonNo ratings yet
- Muscle & Nerve Volume 15 Issue 3 1992 (Doi 10 - CopiarDocument10 pagesMuscle & Nerve Volume 15 Issue 3 1992 (Doi 10 - CopiarPile BallNo ratings yet
- Dampney 2009Document1 pageDampney 2009elbueno21No ratings yet
- Neurochemistry of The Afferents To The Rat Cochlear Root Nucleus: Possible Synaptic Modulation of The Acoustic StartleDocument14 pagesNeurochemistry of The Afferents To The Rat Cochlear Root Nucleus: Possible Synaptic Modulation of The Acoustic StartleJavier HerreroNo ratings yet
- OrlDocument14 pagesOrlchadi_lbNo ratings yet
- Bear&KirkwoodDocument11 pagesBear&KirkwoodswagatarcNo ratings yet
- J Clinph 2005 04 020Document11 pagesJ Clinph 2005 04 020Andres Rojas JerezNo ratings yet
- 1983 - Partial Infraorbital Nerve Ligation As A Model of Trigeminal Nerve Injury in The Mouse Behavioral, NDocument13 pages1983 - Partial Infraorbital Nerve Ligation As A Model of Trigeminal Nerve Injury in The Mouse Behavioral, NNarawut Paka7eprotNo ratings yet
- Leathers&Olson2012-LIP Neurons Encode Cue SalienceDocument5 pagesLeathers&Olson2012-LIP Neurons Encode Cue Salienceiulia andreeaNo ratings yet
- Norepinephrine Modulates Single Hypothalamic Arcuate Neurons ViaDocument12 pagesNorepinephrine Modulates Single Hypothalamic Arcuate Neurons ViaMariann DuzzNo ratings yet
- Inhibition of NR2B Phosphorylation Restores Alterations in NMDA Receptor Expression and Improves Functional Recovery Following Traumatic Brain Injury in Mice (2008)Document13 pagesInhibition of NR2B Phosphorylation Restores Alterations in NMDA Receptor Expression and Improves Functional Recovery Following Traumatic Brain Injury in Mice (2008)Apróné Török IbolyaNo ratings yet
- Art. L Zrinzo - Intra-Operative STN DBS Attenuates The Prominent Beta Rhythm in The STN in Parkinson's DiseaseDocument8 pagesArt. L Zrinzo - Intra-Operative STN DBS Attenuates The Prominent Beta Rhythm in The STN in Parkinson's DiseaseEstereotaxia BrasilNo ratings yet
- NIH Public Access: Author ManuscriptDocument13 pagesNIH Public Access: Author ManuscriptRoar SyltebøNo ratings yet
- Andsberg 1998 EJNDocument11 pagesAndsberg 1998 EJNZaal KokaiaNo ratings yet
- Overview of Intraoperative Neurophysiological Monitoring During Spinal Surgery - 2016 PDFDocument7 pagesOverview of Intraoperative Neurophysiological Monitoring During Spinal Surgery - 2016 PDFAndrea RoesnerNo ratings yet
- Norepinephrine-Deficient Mice Have Increased Susceptibility To Seizure-Inducing StimuliDocument8 pagesNorepinephrine-Deficient Mice Have Increased Susceptibility To Seizure-Inducing StimuliFrontiersNo ratings yet
- Awards Works: Expanded AbstractDocument2 pagesAwards Works: Expanded AbstractJelvis BofNo ratings yet
- Oral Session I Which Theory Do The Structural Brain Changes Support?Document2 pagesOral Session I Which Theory Do The Structural Brain Changes Support?antonioopNo ratings yet
- Microtubule-Associated Protein MAP2 Expression in Olfactory Bulb in SchizophreniaDocument7 pagesMicrotubule-Associated Protein MAP2 Expression in Olfactory Bulb in SchizophreniagiudisNo ratings yet
- Moore 1996Document23 pagesMoore 1996M4shroomNo ratings yet
- TMP 3564Document10 pagesTMP 3564FrontiersNo ratings yet
- 1 s2.0 S1388245700002480 MainDocument4 pages1 s2.0 S1388245700002480 MainMoon PowerNo ratings yet
- Reduced Neuropeptide Concentrations in Suicide Brain: PeterDocument8 pagesReduced Neuropeptide Concentrations in Suicide Brain: PeterRenata ScavoneNo ratings yet
- Schridde2006 IhDocument13 pagesSchridde2006 Ihiulia andreeaNo ratings yet
- WachowiakDocument16 pagesWachowiakGerónimo Medrano LoeraNo ratings yet
- 2914 FullDocument6 pages2914 FullJean Pierre Chastre LuzaNo ratings yet
- NIH Public Access: Author ManuscriptDocument26 pagesNIH Public Access: Author ManuscriptHippokampaNo ratings yet
- Effects of Sleep Deprivation On Spontane PDFDocument2 pagesEffects of Sleep Deprivation On Spontane PDFAleksandar NusicNo ratings yet
- mRNA Expression of Activity-Regulated Cytoskeleton-Associated Protein (Arc) in The Amygdala-Kindled RatsDocument11 pagesmRNA Expression of Activity-Regulated Cytoskeleton-Associated Protein (Arc) in The Amygdala-Kindled RatsCarlos LeeNo ratings yet
- kerasnoudis2014Document10 pageskerasnoudis2014Laura A M MNo ratings yet
- Cerebral Correlates of Delta Waves During non-REM Sleep RevisitedDocument8 pagesCerebral Correlates of Delta Waves During non-REM Sleep RevisitedEmiliano PrandelliNo ratings yet
- Berning 2012Document1 pageBerning 2012László SágiNo ratings yet
- Nikken MX2R Device, Pulsed Magnetic Field Therapy in Refractory CarpalDocument6 pagesNikken MX2R Device, Pulsed Magnetic Field Therapy in Refractory CarpalgstrohlNo ratings yet
- Orexin Neuron Activity in Mating Mice - A Pilot STDocument6 pagesOrexin Neuron Activity in Mating Mice - A Pilot STRika LauraNo ratings yet
- 2003 Changes in Median Nerve Somatosensory Transmission and Motor OutputDocument12 pages2003 Changes in Median Nerve Somatosensory Transmission and Motor OutputchooNo ratings yet
- Oorschot 1996Document20 pagesOorschot 1996M4shroomNo ratings yet
- Jmf.2020.0137-Genoderma in PDDocument8 pagesJmf.2020.0137-Genoderma in PDApostolos T.No ratings yet
- Depression SerotoninDocument5 pagesDepression SerotoninidjacobsNo ratings yet
- PAIN IN PARKINSON'S DISEASE-50 Cases (2008)Document4 pagesPAIN IN PARKINSON'S DISEASE-50 Cases (2008)Apostolos T.No ratings yet
- MiorelDocument1 pageMiorelApostolos T.No ratings yet
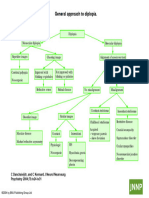
- Diplopia-jnnp-2004-December-75-suppl 4-Iv24Document1 pageDiplopia-jnnp-2004-December-75-suppl 4-Iv24Apostolos T.No ratings yet
- Assessment of Efficacy and Safety of Eslicarbazepine Acetate For The Treatment of Trigeminal NeuralgiaDocument9 pagesAssessment of Efficacy and Safety of Eslicarbazepine Acetate For The Treatment of Trigeminal NeuralgiaApostolos T.No ratings yet
- PAINWeek Journal Vol 5, Q1Document62 pagesPAINWeek Journal Vol 5, Q1Apostolos T.No ratings yet
- Diplopia - Eye Disorders - MSD Manual Professional EditionDocument5 pagesDiplopia - Eye Disorders - MSD Manual Professional EditionApostolos T.No ratings yet
- Vimpat Tablets Product CharacteristicsDocument148 pagesVimpat Tablets Product CharacteristicsApostolos T.No ratings yet
- Diplopia - Eye Disorders - MSD Manual Professional Edition-TableDocument3 pagesDiplopia - Eye Disorders - MSD Manual Professional Edition-TableApostolos T.No ratings yet
- Pain: Current Understanding of Assessment, Management, and TreatmentsDocument101 pagesPain: Current Understanding of Assessment, Management, and TreatmentsNational Pharmaceutical Council100% (8)
- Diagnosing Secondary Headaches (Practical Neurology) (2020 - 05)Document5 pagesDiagnosing Secondary Headaches (Practical Neurology) (2020 - 05)Apostolos T.No ratings yet
- Trigemina Neuralgia Guidelines - 2021 - V4Document21 pagesTrigemina Neuralgia Guidelines - 2021 - V4Apostolos T.No ratings yet
- PAINWeek Journal Vol 7, Q1Document78 pagesPAINWeek Journal Vol 7, Q1Apostolos T.No ratings yet
- CONy 2020 Virtual Congress Scientific Program OverviewDocument24 pagesCONy 2020 Virtual Congress Scientific Program OverviewApostolos T.No ratings yet
- Understanding Cluster Headache Using MRI (2020)Document12 pagesUnderstanding Cluster Headache Using MRI (2020)Apostolos T.No ratings yet
- Fibromyalgia & Trazodone+Pregabalin 2011Document9 pagesFibromyalgia & Trazodone+Pregabalin 2011Apostolos T.No ratings yet
- NIH Public Access: Author ManuscriptDocument48 pagesNIH Public Access: Author ManuscriptApostolos T.No ratings yet
- ITCHDocument18 pagesITCHApostolos T.No ratings yet
- The Status of Diagnosis and Treatment To Intracranial Hypotension (2017)Document8 pagesThe Status of Diagnosis and Treatment To Intracranial Hypotension (2017)Apostolos T.No ratings yet
- Overview of Standardized HRV Assessment Protocol - 2Document7 pagesOverview of Standardized HRV Assessment Protocol - 2Apostolos T.No ratings yet
- Adverse Effects of Antiepileptic Drugs (2010)Document8 pagesAdverse Effects of Antiepileptic Drugs (2010)Apostolos T.No ratings yet
- Neuroimaging in Cluster Headache and Other Trigeminal Autonomic Cephalalgias (2012)Document10 pagesNeuroimaging in Cluster Headache and Other Trigeminal Autonomic Cephalalgias (2012)Apostolos T.No ratings yet
- Cephalalgia ReviewDocument25 pagesCephalalgia ReviewApostolos T.No ratings yet
- Functional Magnetic Resonance Imaging in Episodic Cluster Headache (2009)Document4 pagesFunctional Magnetic Resonance Imaging in Episodic Cluster Headache (2009)Apostolos T.No ratings yet
- Setup: Original Language (English) Your Language TranslationDocument25 pagesSetup: Original Language (English) Your Language TranslationApostolos T.No ratings yet
- Metals: Iron, Aging, and NeurodegenerationDocument23 pagesMetals: Iron, Aging, and NeurodegenerationApostolos T.No ratings yet
- Adult EpilepsyDocument14 pagesAdult Epilepsyammarhafez78No ratings yet
- A Role For FMRI in Optimizing CNS Drug DDocument14 pagesA Role For FMRI in Optimizing CNS Drug DApostolos T.No ratings yet
- Metals: Iron, Aging, and NeurodegenerationDocument23 pagesMetals: Iron, Aging, and NeurodegenerationApostolos T.No ratings yet
- Hydrocarbon ReactionsDocument2 pagesHydrocarbon ReactionsJessa Libo-onNo ratings yet
- A Perspective in Accelerated Orthodontics With Aligner Treatment 2017 Seminars in OrthodonticsDocument7 pagesA Perspective in Accelerated Orthodontics With Aligner Treatment 2017 Seminars in Orthodonticsdruzair007No ratings yet
- 8-Input Multiplexer Selects One of Eight InputsDocument3 pages8-Input Multiplexer Selects One of Eight InputsKang RoyNo ratings yet
- Gamma Ray Spectroscopy ExperimentDocument15 pagesGamma Ray Spectroscopy ExperimentLizelleNiit100% (1)
- Asme Code Sec Ix Ma Appe 2004Document3 pagesAsme Code Sec Ix Ma Appe 2004Guillermo CamachoNo ratings yet
- Chapter11 Part1 PDFDocument40 pagesChapter11 Part1 PDFvampanezezNo ratings yet
- Abstract of SURYA NAMASKARDocument23 pagesAbstract of SURYA NAMASKARSarita SharmaNo ratings yet
- Project On Baggage Handling and AcceptanceDocument52 pagesProject On Baggage Handling and AcceptanceKiran Reddy50% (2)
- VTBS 20-3DDocument1 pageVTBS 20-3Dwong keen faivNo ratings yet
- 12.3 Operation Qualification Protocol For Laminar Air Flow UnitDocument4 pages12.3 Operation Qualification Protocol For Laminar Air Flow UnituzairNo ratings yet
- Hilti Bolt DesignDocument7 pagesHilti Bolt Designcheligp1981No ratings yet
- Rift Vally BC N RegularDocument308 pagesRift Vally BC N RegularCabdisacid yasinNo ratings yet
- Ic F3022SDocument36 pagesIc F3022SEfren.galNo ratings yet
- DTP - Digital Transformer ProtectionDocument13 pagesDTP - Digital Transformer ProtectionHBNBILNo ratings yet
- Taylor Ebooks 4 Web 0218Document360 pagesTaylor Ebooks 4 Web 0218aquahellNo ratings yet
- Q's For CONS VivaDocument251 pagesQ's For CONS VivaHanin AbukhiaraNo ratings yet
- Manual stack emission monitoring standardDocument29 pagesManual stack emission monitoring standardsantisal11No ratings yet
- Phases of Clinical Trials: OverlapDocument3 pagesPhases of Clinical Trials: OverlapMohammed El-GezeiryNo ratings yet
- BA Anchor Data SheetDocument17 pagesBA Anchor Data SheetbrodieNo ratings yet
- On K-Distance Degree Index of TreesDocument5 pagesOn K-Distance Degree Index of TreesVelumani sNo ratings yet
- #Healthy Food Recipes EasyDocument4 pages#Healthy Food Recipes EasyTHE NEXT GENNo ratings yet
- Wound Care, Dressing and BandagingDocument11 pagesWound Care, Dressing and BandagingJessica Febrina Wuisan100% (1)
- Gardening Tool Identification QuizDocument3 pagesGardening Tool Identification Quizcaballes.melchor86No ratings yet
- IV Solution Cheat Sheet: Type Description Osmolality Use MiscellaneousDocument1 pageIV Solution Cheat Sheet: Type Description Osmolality Use MiscellaneousKristine Castillo100% (2)
- Nexon Genuine AccessoriesDocument15 pagesNexon Genuine AccessoriesjagaaniNo ratings yet
- TSC-2015-TAS Dec 10 PDFDocument8 pagesTSC-2015-TAS Dec 10 PDFvempadareddyNo ratings yet
- Hoist by TechnologyDocument6 pagesHoist by TechnologyAnonymous ntE0hG2TPNo ratings yet
- Life Saving Appliance: Personal Life-Saving Appliances Lifeboats & Rescue Boats LiferaftsDocument18 pagesLife Saving Appliance: Personal Life-Saving Appliances Lifeboats & Rescue Boats Liferaftsdafa dzaky100% (1)
- The Volcanic Explosivity Index (VEI)Document8 pagesThe Volcanic Explosivity Index (VEI)Rakhmatul ArafatNo ratings yet
- Effect of Blast Induced Vibrations On GroutDocument55 pagesEffect of Blast Induced Vibrations On GroutDavid marpaungNo ratings yet