You might also like
- My Genealogy Story Part 5: New DiscoveriesDocument65 pagesMy Genealogy Story Part 5: New DiscoveriesTimothyNo ratings yet
- Mendelian Genetics Worksheet PDFDocument4 pagesMendelian Genetics Worksheet PDFgil100% (1)
- The 14 Healing Plexus PDFDocument6 pagesThe 14 Healing Plexus PDFSaravanan Sivasangaran100% (1)
- Odum Fundamentals of EcologyDocument17 pagesOdum Fundamentals of Ecologyelizstrov64% (45)
- Mitacs Accelerate Proposal TemplateDocument18 pagesMitacs Accelerate Proposal TemplatenadhemNo ratings yet
- Flow Patterns in The Radiocephalic Arteriovenous "Stula: An in Vitro StudyDocument11 pagesFlow Patterns in The Radiocephalic Arteriovenous "Stula: An in Vitro StudyJenny GoNo ratings yet
- 1 s2.0 0735109788900940 MainDocument11 pages1 s2.0 0735109788900940 MainHarish SubramanianNo ratings yet
- From Tile Department of Anatomy, School of Medicine, University of California, and Veterans Administration Center, Los AngelesDocument15 pagesFrom Tile Department of Anatomy, School of Medicine, University of California, and Veterans Administration Center, Los Angelesarclite86No ratings yet
- Human: The Microscopic Structure Normal Synovial TissueDocument12 pagesHuman: The Microscopic Structure Normal Synovial TissueKENZA EL ALAOUINo ratings yet
- Sala The 1976Document23 pagesSala The 1976nitesh_kumar079976No ratings yet
- Energy Harvesting Eel: JFLS 20000355Document13 pagesEnergy Harvesting Eel: JFLS 20000355hextobinaryNo ratings yet
- Abdominal Aortic AneurysmDocument23 pagesAbdominal Aortic AneurysmHani AbulkhairNo ratings yet
- Biophysj00661 0041Document16 pagesBiophysj00661 0041jimNo ratings yet
- A Study On Two-Layered Model (Casson-Newtonian) For Blood Ow Through An Arterial Stenosis: Axially Variable Slip Velocity at The WallDocument14 pagesA Study On Two-Layered Model (Casson-Newtonian) For Blood Ow Through An Arterial Stenosis: Axially Variable Slip Velocity at The WallRicardoNo ratings yet
- Centrel Venous CatheterizationDocument77 pagesCentrel Venous CatheterizationAli100% (1)
- Anatomy: Initiation Factors Important To Which Are Believed To Be Axial StreamDocument11 pagesAnatomy: Initiation Factors Important To Which Are Believed To Be Axial StreamogangurelNo ratings yet
- Examination of The Soft Tissues of The ChestDocument8 pagesExamination of The Soft Tissues of The ChestDum MyNo ratings yet
- Efectos de La Parotina en Huesos LargosDocument6 pagesEfectos de La Parotina en Huesos LargosAlejandro GuzmanNo ratings yet
- Arthrography of Shoulder PDFDocument12 pagesArthrography of Shoulder PDFTanya GoriNo ratings yet
- A Sella Turcica Bridge in Subjects With Severe Craniofacial DeviationsDocument6 pagesA Sella Turcica Bridge in Subjects With Severe Craniofacial DeviationskalixinNo ratings yet
- Of Physiology, College, London,: 445 Text-IiguremDocument13 pagesOf Physiology, College, London,: 445 Text-IiguremDejan ZikicNo ratings yet
- Vibrations of Orthotropic Sandwich Conical Shells With Free EdgesDocument14 pagesVibrations of Orthotropic Sandwich Conical Shells With Free EdgesunlockvnNo ratings yet
- Torneck Part IDocument9 pagesTorneck Part IEfren FloresNo ratings yet
- Blood Supply To The Maxillary SinusDocument12 pagesBlood Supply To The Maxillary Sinusprostfang100% (2)
- 10 1007@BF01276692Document11 pages10 1007@BF01276692Pilar AufrastoNo ratings yet
- CFD Simulation of Carotid Artery Stenosis - Simplified ModelDocument16 pagesCFD Simulation of Carotid Artery Stenosis - Simplified ModelMolla NayonNo ratings yet
- FksmJMedSci 50 p47Document10 pagesFksmJMedSci 50 p47Abhilash VemulaNo ratings yet
- Pie AnalisisDocument4 pagesPie AnalisisniorNo ratings yet
- Complex-Periodic Spiral Waves in Confluent Cardiac Cell Cultures Induced by Localized InhomogeneitiesDocument7 pagesComplex-Periodic Spiral Waves in Confluent Cardiac Cell Cultures Induced by Localized InhomogeneitiesGretymjNo ratings yet
- Heel Spur Formation and The Subcalcaneal Enthesis of The Plantar FasciaDocument3 pagesHeel Spur Formation and The Subcalcaneal Enthesis of The Plantar FasciaanurNo ratings yet
- Fujiwara 2009Document6 pagesFujiwara 2009UserNo ratings yet
- Is Airway Closure CausedDocument16 pagesIs Airway Closure CausedfaberNo ratings yet
- PIIS0022202X15449565Document8 pagesPIIS0022202X15449565Novrina Wahidah RestiNo ratings yet
- Tortuosity,: Coiling, and Kinking of Internal Carotid ArteryDocument13 pagesTortuosity,: Coiling, and Kinking of Internal Carotid ArteryRoberto AmayaNo ratings yet
- The Sarcomere Length-Tension Relation in Skeletal MuscleDocument28 pagesThe Sarcomere Length-Tension Relation in Skeletal MuscleDanielle GraceNo ratings yet
- !5uroical 5ection.: Some Points Surgical of Supply Hamilton Drummond, F.R.C.S.EdDocument9 pages!5uroical 5ection.: Some Points Surgical of Supply Hamilton Drummond, F.R.C.S.EdaliNo ratings yet
- Evaluation of Fatigue Models - Nigerian StandardDocument12 pagesEvaluation of Fatigue Models - Nigerian StandardDATEME ABAMNo ratings yet
- General Surgery Загальна хірургія - 2023 - № 2 (5)Document8 pagesGeneral Surgery Загальна хірургія - 2023 - № 2 (5)Pavlo BadiulNo ratings yet
- Neuralgia MandibularDocument4 pagesNeuralgia Mandibularkubra_tuncel6296No ratings yet
- Artigo 01Document14 pagesArtigo 01Luke OliveiraNo ratings yet
- On Stationary and Travelling Vortex Breakdowns: Byturgut SarpkayaDocument24 pagesOn Stationary and Travelling Vortex Breakdowns: Byturgut Sarpkayaamitchawla1095No ratings yet
- Al-Kayat and BramliDocument13 pagesAl-Kayat and BramliAshish Dadhania100% (1)
- Reduction IN Bronchial Subdivision BronchiectasisDocument15 pagesReduction IN Bronchial Subdivision BronchiectasisyikuiNo ratings yet
- Two-Dimensional Blood #Ow Through Tapered Arteries Under Stenotic ConditionsDocument15 pagesTwo-Dimensional Blood #Ow Through Tapered Arteries Under Stenotic ConditionsAmanullah KabirNo ratings yet
- Gambar Usg TestisDocument28 pagesGambar Usg TestisIrfan YHNo ratings yet
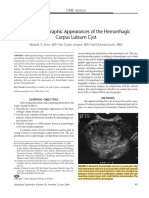
- Various Sonographic Appearances of The Hemorrhagic Corpus Luteum CystDocument14 pagesVarious Sonographic Appearances of The Hemorrhagic Corpus Luteum CystBerry BancinNo ratings yet
- Glaucoma Glaucoma Surgery: Section 2Document62 pagesGlaucoma Glaucoma Surgery: Section 2Imran SyedNo ratings yet
- Fundamental Considerations of The Design and Function of Intranasal AntrostomiesDocument4 pagesFundamental Considerations of The Design and Function of Intranasal AntrostomiessevattapillaiNo ratings yet
- Journal of Entomology and Zoology Volume Eleven, Number Two, June 1919From EverandJournal of Entomology and Zoology Volume Eleven, Number Two, June 1919No ratings yet
- Bogduk Meniscoide PDFDocument15 pagesBogduk Meniscoide PDFMatias Hassan LattzNo ratings yet
- Cole 1926Document39 pagesCole 1926J.D. NobleNo ratings yet
- CT Scan of The AxillaDocument28 pagesCT Scan of The AxillaObamaNo ratings yet
- Kim1985 PDFDocument7 pagesKim1985 PDFfadwa lachkerNo ratings yet
- River MeandersDocument25 pagesRiver MeandersgeoecologistNo ratings yet
- Clinical Anatomy of The Maxillary Artery: Ippei OTAKE, Ikuo Kageyama and Izumi MATAGADocument10 pagesClinical Anatomy of The Maxillary Artery: Ippei OTAKE, Ikuo Kageyama and Izumi MATAGARajan KarmakarNo ratings yet
- Increasing Clamping ForcesDocument7 pagesIncreasing Clamping ForcesAyhan ComertNo ratings yet
- 705 FullDocument9 pages705 FullMauricio SchneiderNo ratings yet
- Cancer - October 1976 - Fishkin - Cervical Lymph Node Metastasis As The First Manifestation of Localized ExtramedullaryDocument4 pagesCancer - October 1976 - Fishkin - Cervical Lymph Node Metastasis As The First Manifestation of Localized ExtramedullarymacklynmanuelNo ratings yet
- Will The Real Ventricular Architecture Please StanDocument13 pagesWill The Real Ventricular Architecture Please Stannilberto2No ratings yet
- Broen 1964Document9 pagesBroen 1964Min ThuNo ratings yet
- Bast 1928Document5 pagesBast 1928silviaNo ratings yet
- Luthi 81 Experiments On Non Channelized Turbidity Currents and Their DepositsDocument10 pagesLuthi 81 Experiments On Non Channelized Turbidity Currents and Their DepositsKe YamNo ratings yet
- Lec. 1 Selected TopicsDocument30 pagesLec. 1 Selected TopicsMustafa KamelNo ratings yet
- Electrophoresis in Stabilizing Media: Paper Chromatography and ElectrophoresisFrom EverandElectrophoresis in Stabilizing Media: Paper Chromatography and ElectrophoresisNo ratings yet
- Da Costa 1903Document7 pagesDa Costa 1903Pilar AufrastoNo ratings yet
- 1 s2.0 S0375960109000590 MainDocument5 pages1 s2.0 S0375960109000590 MainPilar AufrastoNo ratings yet
- Spatio-Temporal Evolution of Extracellular Electric Potential Upon Starvation Around Plasmodia of Physarum-PolycephalumDocument16 pagesSpatio-Temporal Evolution of Extracellular Electric Potential Upon Starvation Around Plasmodia of Physarum-PolycephalumPilar AufrastoNo ratings yet
- Campbell 1992Document3 pagesCampbell 1992Pilar AufrastoNo ratings yet
- Jaworowski 1999 Radiation Risk and EthicsDocument7 pagesJaworowski 1999 Radiation Risk and EthicsPilar AufrastoNo ratings yet
- Pedchenko 2021Document9 pagesPedchenko 2021Pilar AufrastoNo ratings yet
- Fema 2023Document258 pagesFema 2023Pilar AufrastoNo ratings yet
- Adenosine Role in CancerDocument21 pagesAdenosine Role in CancerPilar AufrastoNo ratings yet
- Hanahan 2022Document16 pagesHanahan 2022Pilar AufrastoNo ratings yet
- Li 2019 - Association of Postdisaster Depression and Posttraumatic Stress Disorder With Mortality Among Older Disaster Survivors of The 2011 Great East Japan Earthquake and TsunamiDocument12 pagesLi 2019 - Association of Postdisaster Depression and Posttraumatic Stress Disorder With Mortality Among Older Disaster Survivors of The 2011 Great East Japan Earthquake and TsunamiPilar AufrastoNo ratings yet
- Steinbock - Reaction-Diffusion Model To Solve A Maze - Science 1995Document5 pagesSteinbock - Reaction-Diffusion Model To Solve A Maze - Science 1995Pilar AufrastoNo ratings yet
- Hick 2017 Proposed-Exposure-And-Symptom-Triage-East-Tool-To-Assess-Radiation-Exposure-After-A-Nuclear-DetonationDocument10 pagesHick 2017 Proposed-Exposure-And-Symptom-Triage-East-Tool-To-Assess-Radiation-Exposure-After-A-Nuclear-DetonationPilar AufrastoNo ratings yet
- Guan 2023 Anim Models and Exp Med - Development of Radiation Countermeasure Agents For Acute Radiation SyndromesDocument8 pagesGuan 2023 Anim Models and Exp Med - Development of Radiation Countermeasure Agents For Acute Radiation SyndromesPilar AufrastoNo ratings yet
- Kok 2020Document32 pagesKok 2020Pilar AufrastoNo ratings yet
- Bomanji 2014 Radiation Accidents and Their Management - Role of Medicine ProfessionalsDocument8 pagesBomanji 2014 Radiation Accidents and Their Management - Role of Medicine ProfessionalsPilar AufrastoNo ratings yet
- Sharma 2023 - Nature - Assembly Theory Explains and Quantifies Natural Selection and EvolutionDocument12 pagesSharma 2023 - Nature - Assembly Theory Explains and Quantifies Natural Selection and EvolutionPilar AufrastoNo ratings yet
- Sloane1981 - Science - CathepsinsDocument4 pagesSloane1981 - Science - CathepsinsPilar AufrastoNo ratings yet
- Scrotal CancerDocument12 pagesScrotal CancerPilar AufrastoNo ratings yet
- Santana Codina Et Al 2017 The Role of Autophagy in CancerDocument23 pagesSantana Codina Et Al 2017 The Role of Autophagy in CancerPilar AufrastoNo ratings yet
- Fenno (2005), Laboratory Maintenance of Treponema DenticolaDocument21 pagesFenno (2005), Laboratory Maintenance of Treponema DenticolaPhuong ThaoNo ratings yet
- The Death of The Moth: By: Virginia WoolfDocument8 pagesThe Death of The Moth: By: Virginia WoolfCrizelda CardenasNo ratings yet
- Chap 3 - Group 1 - The Cambrian Explosion and Protistan AncestryDocument26 pagesChap 3 - Group 1 - The Cambrian Explosion and Protistan AncestryRachel TomlayenNo ratings yet
- 1 Compressed Notes BioDDocument23 pages1 Compressed Notes BioDLIM ZHI SHUENNo ratings yet
- Moist Heat Sterlization Liq.Document6 pagesMoist Heat Sterlization Liq.Dr usama El ShafeyNo ratings yet
- PENGARUH PH TERHADAP AKTIVITAS ENZIMDocument14 pagesPENGARUH PH TERHADAP AKTIVITAS ENZIMAco LannaNo ratings yet
- Archives of Biochemistry and BiophysicsDocument10 pagesArchives of Biochemistry and BiophysicsJuanNo ratings yet
- Introductory Notes On Pro Life ApologeticsDocument30 pagesIntroductory Notes On Pro Life ApologeticskedronjNo ratings yet
- DIASS NotesDocument8 pagesDIASS NotesBern Patrick BautistaNo ratings yet
- Biotech L1 Cell Transport L2 Cell Division 1Document18 pagesBiotech L1 Cell Transport L2 Cell Division 1ColourBlueNo ratings yet
- General Biology 102 Laboratory: Quiz #2 (20 Points)Document1 pageGeneral Biology 102 Laboratory: Quiz #2 (20 Points)obnewullNo ratings yet
- Lamarckism and Neo LamarckismDocument5 pagesLamarckism and Neo LamarckismImran HussainNo ratings yet
- Identification and Collection of Macro Fungi (Basidiomycota: Agaricales) From Baturraden Botanical Garden, BanyumasDocument5 pagesIdentification and Collection of Macro Fungi (Basidiomycota: Agaricales) From Baturraden Botanical Garden, BanyumaslutfianaNo ratings yet
- Ap Lab Mitosis CancerDocument11 pagesAp Lab Mitosis Cancerapi-38237256450% (6)
- The Psychogenesis of Knowledge and Its Epistemological SignificanceDocument4 pagesThe Psychogenesis of Knowledge and Its Epistemological SignificanceGil CostaNo ratings yet
- The IQ of Gypsies in Central EuropeDocument11 pagesThe IQ of Gypsies in Central EuropeAlexandru CiorobescuNo ratings yet
- Shenzhen CodeDocument293 pagesShenzhen CodeAnabela PlosNo ratings yet
- Bsa ProspectusDocument2 pagesBsa ProspectusKeziah Millones100% (1)
- PERIPHERAL SMEAR - UG ClassDocument71 pagesPERIPHERAL SMEAR - UG Classaditya.3757No ratings yet
- EVOLUTION-By Dr. Sunita SaxenaDocument172 pagesEVOLUTION-By Dr. Sunita SaxenaDivya AgarawalNo ratings yet
- Leaf Concept MapDocument1 pageLeaf Concept MapGraceAbarcaNo ratings yet
- Ecological RelationshipDocument2 pagesEcological RelationshipRochel LimNo ratings yet
- Experiment 4: Isolation of Microbes From Environmental SampleDocument3 pagesExperiment 4: Isolation of Microbes From Environmental SampleAnis SurayaNo ratings yet
- Solutions For Practice Problems For Molecular Biology, Session 2: DNA ReplicationDocument4 pagesSolutions For Practice Problems For Molecular Biology, Session 2: DNA ReplicationRahmadyanTefaraniNo ratings yet
- Exploring Science JC Sample PagesDocument18 pagesExploring Science JC Sample Pagesmiek ThomasNo ratings yet