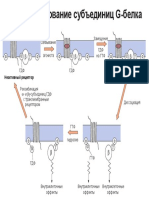
You might also like
- The Emerging Physiological Roles of The Glycerophosphodiesterase FamilyDocument19 pagesThe Emerging Physiological Roles of The Glycerophosphodiesterase FamilyArshia NazirNo ratings yet
- 5 Substrate Selectivity in Photoreceptors: RGS9-GDocument9 pages5 Substrate Selectivity in Photoreceptors: RGS9-GscotNo ratings yet
- CD2BP2-GYF Interacts With The Spliceosomal Protein SMB - B' 1 (PDFDrive)Document31 pagesCD2BP2-GYF Interacts With The Spliceosomal Protein SMB - B' 1 (PDFDrive)Blink SNo ratings yet
- GPCR Review - Lefkovitz - NatureRev - 2002Document12 pagesGPCR Review - Lefkovitz - NatureRev - 2002EvaNo ratings yet
- Regulation of Phospholipase C by G Proteins: Bordatella PertussisDocument5 pagesRegulation of Phospholipase C by G Proteins: Bordatella Pertussissant_grNo ratings yet
- PIIS1535947620300542Document15 pagesPIIS1535947620300542Blink SNo ratings yet
- Syrovatkina Et Al. - 2016 - Regulation, Signaling, And Physiological FunctionsDocument40 pagesSyrovatkina Et Al. - 2016 - Regulation, Signaling, And Physiological Functionskdh03081995No ratings yet
- AGP Review: Structure, Expression and FunctionsDocument15 pagesAGP Review: Structure, Expression and Functionsmahmoud korieshNo ratings yet
- SepiaDocument10 pagesSepiaAmaliaNo ratings yet
- Targeted Disruption of The GlucocorticoidDocument15 pagesTargeted Disruption of The GlucocorticoidSyifa SariNo ratings yet
- Casas 2017Document10 pagesCasas 2017Margarita SantoyoNo ratings yet
- For Isha GPCRsDocument23 pagesFor Isha GPCRsSALEHA HASSANNo ratings yet
- Superfamily of G-Protein Coupled Receptors (GPCRS) - Extraordinary and Outstanding Success of EvolutionDocument13 pagesSuperfamily of G-Protein Coupled Receptors (GPCRS) - Extraordinary and Outstanding Success of EvolutionSELNo ratings yet
- Multiplex GPCR Screen Reveals Reliably Acting Agonists and A Gq-Phospholipase C Coupling Mode of GPR30/GPER1Document57 pagesMultiplex GPCR Screen Reveals Reliably Acting Agonists and A Gq-Phospholipase C Coupling Mode of GPR30/GPER1Benjamin VillagraNo ratings yet
- Paiardini Et Al-2018-Proteins: Structure, Function, and Bioinformatics PDFDocument27 pagesPaiardini Et Al-2018-Proteins: Structure, Function, and Bioinformatics PDFVignesh NarayanNo ratings yet
- Molecular Basis of Gephyrin Clustering at Inhibitory SynapsesDocument8 pagesMolecular Basis of Gephyrin Clustering at Inhibitory SynapsesAndor SzekeresNo ratings yet
- Daaka 2012Document9 pagesDaaka 2012iara rocha xavierNo ratings yet
- N-Acetyl-: - Glucosaminylphosphatidylinositol De-N-acetylase From Entamoeba HistolyticaDocument7 pagesN-Acetyl-: - Glucosaminylphosphatidylinositol De-N-acetylase From Entamoeba HistolyticaDr. Mohd Ashraf KhanNo ratings yet
- Cordier - Mendes Et Al. 2007 - A Metabolic and Genomic Study of Engineered Saccharomyces Cerevisiae Strains For High Glycerol ProductionDocument15 pagesCordier - Mendes Et Al. 2007 - A Metabolic and Genomic Study of Engineered Saccharomyces Cerevisiae Strains For High Glycerol ProductionBrian VasquezNo ratings yet
- G-Protein Coupled Receptors and Their Signaling MechanismDocument21 pagesG-Protein Coupled Receptors and Their Signaling Mechanismaroob9mahamoodaurinNo ratings yet
- Screening of Peptide Ligands For Pyrroloquinoline Quinone Glucose Dehydrogenase Using Antagonistic Template-Based BiopanningDocument13 pagesScreening of Peptide Ligands For Pyrroloquinoline Quinone Glucose Dehydrogenase Using Antagonistic Template-Based BiopanningGabriel CesarNo ratings yet
- RGD Peptides Product BrochureDocument8 pagesRGD Peptides Product BrochureRobert BrousseauNo ratings yet
- Mini GDocument8 pagesMini GOlivia LuciannoNo ratings yet
- Ikemoto 2002Document13 pagesIkemoto 2002cataawwwNo ratings yet
- Types of Signalling Pathway GPCR Pathway Normal and DysregulatedDocument78 pagesTypes of Signalling Pathway GPCR Pathway Normal and Dysregulatedapi-251915360No ratings yet
- G - Protein Coupled ReceptersDocument20 pagesG - Protein Coupled ReceptersShaikh AmeeruddinNo ratings yet
- Glutathione Peroxidase Family - An Evolutionary OverviewDocument12 pagesGlutathione Peroxidase Family - An Evolutionary OverviewRogerio MargisNo ratings yet
- G Protein ReviewDocument17 pagesG Protein ReviewSaray Gordián CastroNo ratings yet
- PIIS0021925820816445Document10 pagesPIIS0021925820816445tazira syakinNo ratings yet
- Describe The Signalling Pathways Downstream of The Heterotrimeric G Proteins GS, Gi and GQDocument6 pagesDescribe The Signalling Pathways Downstream of The Heterotrimeric G Proteins GS, Gi and GQwavezone113100% (2)
- Bretthauer Et Al 2009Document6 pagesBretthauer Et Al 2009Marcell CrispimNo ratings yet
- GPCRs Regulate Physiological FunctionsDocument25 pagesGPCRs Regulate Physiological FunctionsMirza Shaharyar BaigNo ratings yet
- Gycosaminoglycans 21Document14 pagesGycosaminoglycans 21Ertugrul Ghazi LegendNo ratings yet
- MIT7 - 05S20 - Pset8 BIOCHEMDocument9 pagesMIT7 - 05S20 - Pset8 BIOCHEMRejoice chekesaNo ratings yet
- Use of Functionalized Poly (Ethylene Glycol) S For Modification of PolypeptidesDocument24 pagesUse of Functionalized Poly (Ethylene Glycol) S For Modification of PolypeptidesFranciscoNo ratings yet
- Janetopoulos Science2001Document5 pagesJanetopoulos Science2001Psych PsychhNo ratings yet
- Prova SuficiênciaDocument4 pagesProva SuficiênciaevilbioNo ratings yet
- G-protein-Coupled ReceptorsDocument24 pagesG-protein-Coupled ReceptorsNaimi Amalia hatimahNo ratings yet
- Red Cell: Blood GlycophorinsDocument11 pagesRed Cell: Blood GlycophorinsChu NgọcNo ratings yet
- Plant Science: The Two Faces of DJ-1D ProteinsDocument3 pagesPlant Science: The Two Faces of DJ-1D ProteinsSahilu RabiluNo ratings yet
- +jianxia Li 2016Document13 pages+jianxia Li 2016ramiroNo ratings yet
- Glutamate Synthases From Conifers, Gene Structure and Phylogenetic Studies BMCDocument12 pagesGlutamate Synthases From Conifers, Gene Structure and Phylogenetic Studies BMCAngel GarciaNo ratings yet
- 1 s2.0 S0006349522008566 MainDocument13 pages1 s2.0 S0006349522008566 MainmanelmanoNo ratings yet
- G-Protein Coupled Receptors: Biogenic Amines Amino Acids and Ions: Lipids: Peptides and ProteinsDocument4 pagesG-Protein Coupled Receptors: Biogenic Amines Amino Acids and Ions: Lipids: Peptides and ProteinsPranav NakhateNo ratings yet
- Crystal Structures of B-Galactosidase From Penicillium Sp. and Its Complex With GalactoseDocument12 pagesCrystal Structures of B-Galactosidase From Penicillium Sp. and Its Complex With GalactoseTruyen MaiNo ratings yet
- Recombinant Expression, in Vitro Refolding, and Biophysical Characterization of The Human Glucagon-Like Peptide-1 ReceptorDocument10 pagesRecombinant Expression, in Vitro Refolding, and Biophysical Characterization of The Human Glucagon-Like Peptide-1 ReceptorDhul FiqarNo ratings yet
- Second Messenger Systems: Figs 1.4 1.5Document1 pageSecond Messenger Systems: Figs 1.4 1.5AntonPurpurovNo ratings yet
- Prostaglandin Synthases: Molecular Characterization and Involvement in Prostaglandin BiosynthesisDocument20 pagesProstaglandin Synthases: Molecular Characterization and Involvement in Prostaglandin BiosynthesisNdayishimiye ErnestNo ratings yet
- Erythrocyte G6PD Activity and GSH Level As Risk Factors For Vascular Complications Among Type 2 Diabetics in Osogbo NigeriaDocument6 pagesErythrocyte G6PD Activity and GSH Level As Risk Factors For Vascular Complications Among Type 2 Diabetics in Osogbo NigeriaAudrey LyliNo ratings yet
- PDGF D and Renal Disease Yet Another One of Those.35Document2 pagesPDGF D and Renal Disease Yet Another One of Those.35Arista RachmaNo ratings yet
- GM3 GangliosideDocument6 pagesGM3 Gangliosideanon_818654990No ratings yet
- Yoshida2003-Phospho of B-Tubulin by GCPRDocument10 pagesYoshida2003-Phospho of B-Tubulin by GCPR9longerNo ratings yet
- Pga DLSJ ReviewDocument6 pagesPga DLSJ ReviewjenniNo ratings yet
- Metabotropic: The Second Major Type of Neurotransmitter ReceptorDocument17 pagesMetabotropic: The Second Major Type of Neurotransmitter ReceptorChian WrightNo ratings yet
- Second Messenger Generation and Destruction: OCTOBER 1992Document5 pagesSecond Messenger Generation and Destruction: OCTOBER 1992sant_grNo ratings yet
- Orphan Receptors 3Document27 pagesOrphan Receptors 3Alonso SánchezNo ratings yet
- Expression and Characteristic of Synthetic Human Epidermal Growth Factor (hEGF) in Transgenic Tobacco PlantsDocument7 pagesExpression and Characteristic of Synthetic Human Epidermal Growth Factor (hEGF) in Transgenic Tobacco PlantsCarlos Julio Nova LopezNo ratings yet
- Covalent Conjugation of Poly (Ethylene Glycol) To Proteins and Peptides: Strategies and MethodsDocument35 pagesCovalent Conjugation of Poly (Ethylene Glycol) To Proteins and Peptides: Strategies and MethodsFranciscoNo ratings yet
- 14 3 3 AssignmentDocument15 pages14 3 3 AssignmentArshia NazirNo ratings yet
- Inclusion Bodies Preparation in E.coliDocument2 pagesInclusion Bodies Preparation in E.coliArshia NazirNo ratings yet
- Enzymes: Principles and Biotechnological ApplicationsDocument41 pagesEnzymes: Principles and Biotechnological ApplicationsDaniela QuiñonesNo ratings yet
- Phases of Adaptive Immune ResponseDocument46 pagesPhases of Adaptive Immune ResponseArshia NazirNo ratings yet
- 1Document5 pages1kunal sharmaNo ratings yet
- FTIR IntroductionDocument8 pagesFTIR Introductiontimtailieu2010No ratings yet
- Duplex PCR For MSSA and MRSADocument3 pagesDuplex PCR For MSSA and MRSAArshia NazirNo ratings yet
- Protein/gzr 048Document12 pagesProtein/gzr 048Arshia NazirNo ratings yet
- Skim Milk Agar: CompositionDocument2 pagesSkim Milk Agar: CompositionArshia NazirNo ratings yet
- Phototactic Behaviour of The Archaebacterial: Natronobacterium PharaonisDocument3 pagesPhototactic Behaviour of The Archaebacterial: Natronobacterium PharaonisArshia NazirNo ratings yet
- Resistance To GentamicinDocument7 pagesResistance To GentamicinArshia NazirNo ratings yet
- Vitek 2 System Principle PDFDocument5 pagesVitek 2 System Principle PDFArshia NazirNo ratings yet
- Aqueous Extract PreparationDocument3 pagesAqueous Extract PreparationArshia NazirNo ratings yet
- Thumb Impression TestDocument11 pagesThumb Impression TestArshia NazirNo ratings yet
- Total Fat Content PDFDocument5 pagesTotal Fat Content PDFArshia NazirNo ratings yet
- FTIR MethodologyDocument12 pagesFTIR MethodologyArshia NazirNo ratings yet
- History and Development of Antimicrobial Susceptibility Testing MethodologyDocument7 pagesHistory and Development of Antimicrobial Susceptibility Testing MethodologyTogu NaiposposNo ratings yet
- DNA of Staph PDFDocument8 pagesDNA of Staph PDFArshia NazirNo ratings yet
- Extra Journal Med TechDocument6 pagesExtra Journal Med TechJeks VSNo ratings yet
- Docking of ProteinDocument14 pagesDocking of ProteinArshia NazirNo ratings yet
- 1 s2.0 S1319610311002407 MainDocument14 pages1 s2.0 S1319610311002407 Mainspanu oanaNo ratings yet
- B Lactamase Behind Methicillin ResistanceDocument2 pagesB Lactamase Behind Methicillin ResistanceArshia NazirNo ratings yet
- Thermus Medium For Storage of T. Aquaticus PDFDocument1 pageThermus Medium For Storage of T. Aquaticus PDFArshia NazirNo ratings yet
- Family FabaceaeDocument10 pagesFamily FabaceaeArshia NazirNo ratings yet
- Review Bauer ArticleDocument6 pagesReview Bauer ArticleArshia NazirNo ratings yet
- Chapter 2 Determination of MICs 2006updated PDFDocument19 pagesChapter 2 Determination of MICs 2006updated PDFRuro LaudaNo ratings yet
- Duabanga Grandiflora (Lythraceae) Against Sitophilus OryzaeDocument6 pagesDuabanga Grandiflora (Lythraceae) Against Sitophilus OryzaeArshia NazirNo ratings yet
- Resazurin As Redox Indicator PDFDocument8 pagesResazurin As Redox Indicator PDFArshia NazirNo ratings yet
- HPLC MethodologyDocument4 pagesHPLC MethodologyArshia NazirNo ratings yet
- Hematologic Tests 3Document5 pagesHematologic Tests 3Sareene Joyce PepitoNo ratings yet
- Test Bank For Foundations of Behavioral Neuroscience Paper 9 e 9th Edition Neil R CarlsonDocument36 pagesTest Bank For Foundations of Behavioral Neuroscience Paper 9 e 9th Edition Neil R Carlsonridenengagedcjeooq100% (45)
- Chapter 6 Reading GuideDocument4 pagesChapter 6 Reading GuideLaila DixonNo ratings yet
- ATP/ADP Energy Cycle ExplainedDocument3 pagesATP/ADP Energy Cycle ExplainedJane Marry IgnacioNo ratings yet
- Laboratory Activity No. 10: Frog Circulatory and Excretory SystemDocument5 pagesLaboratory Activity No. 10: Frog Circulatory and Excretory SystemMaricris GuillermoNo ratings yet
- How cognition shapes affect into emotionDocument21 pagesHow cognition shapes affect into emotionCristina ValenteNo ratings yet
- Virulence Factors of Periodontal PathogensDocument31 pagesVirulence Factors of Periodontal Pathogenst sNo ratings yet
- DUSTIN NUESTRO Skeletal System Lab. WorksheetDocument10 pagesDUSTIN NUESTRO Skeletal System Lab. WorksheetDustin NuestroNo ratings yet
- Care of The Profesional VoiceDocument24 pagesCare of The Profesional VoiceLaura RakauskaitėNo ratings yet
- Prof. N. Syabbalo: MB., CHB., PHD., FCCP., FRS., Fiba Professor of Physiology & MedicineDocument50 pagesProf. N. Syabbalo: MB., CHB., PHD., FCCP., FRS., Fiba Professor of Physiology & MedicineHomeground entertainmentNo ratings yet
- Case Presentation On Myocardial Infarction LalanDocument8 pagesCase Presentation On Myocardial Infarction Lalananimesh pandaNo ratings yet
- Biochemistry Exam 2 (Outline)Document9 pagesBiochemistry Exam 2 (Outline)stanly sotoNo ratings yet
- HUL261: Introduction To Psychology: Altered State of ConsciousnessDocument6 pagesHUL261: Introduction To Psychology: Altered State of ConsciousnessEkant YadavNo ratings yet
- Levin (1993) - Drowning and Near Drowning (Klasifikasi Tenggelam)Document16 pagesLevin (1993) - Drowning and Near Drowning (Klasifikasi Tenggelam)Filbert TandeanNo ratings yet
- 3 PracticalDocument47 pages3 PracticalT NNo ratings yet
- Test Bank For Clinical Application of Mechanical Ventilation 4th Edition Chang Isbn 10 1111539588 Isbn 13 9781111539580Document6 pagesTest Bank For Clinical Application of Mechanical Ventilation 4th Edition Chang Isbn 10 1111539588 Isbn 13 9781111539580Reed Nuss100% (35)
- BlooodDocument14 pagesBlooodMarija JurakovićNo ratings yet
- Finals 2nd Quiz Biochem MALANADocument3 pagesFinals 2nd Quiz Biochem MALANAJay-Ar MalanaNo ratings yet
- Etas Mcqs 2019 PlusDocument1,569 pagesEtas Mcqs 2019 PlusSally dossNo ratings yet
- Alzheimers Disease and Chediak Hijashi SyndromeDocument47 pagesAlzheimers Disease and Chediak Hijashi Syndromecarmelo aguirreNo ratings yet
- Haematological Profile of Severe Acute Malnourished Children Admitted at Our InstitutionDocument4 pagesHaematological Profile of Severe Acute Malnourished Children Admitted at Our InstitutionIJAR JOURNALNo ratings yet
- Unit 7 Urinary SystemDocument5 pagesUnit 7 Urinary SystemSiraj ShiferawNo ratings yet
- 3-Ascending Tracts of Spinal CordDocument28 pages3-Ascending Tracts of Spinal Cordmuhammad altaf100% (2)
- WBC Count: RBC Count, HB, HCT, Blood Indices WBC Count & PLT CountDocument36 pagesWBC Count: RBC Count, HB, HCT, Blood Indices WBC Count & PLT CountppcarpediemNo ratings yet
- NCM 108 - Nursing Care During Normal Pregnancy and Care of The Developing FetusDocument5 pagesNCM 108 - Nursing Care During Normal Pregnancy and Care of The Developing FetusKristine BartsNo ratings yet
- Experiment and Activity CellDocument4 pagesExperiment and Activity CellPang ChixxNo ratings yet
- NCP For Diabetic KetoacidosisDocument2 pagesNCP For Diabetic KetoacidosisLovely Cacapit100% (1)
- Anatomy Central Nervous SystemDocument87 pagesAnatomy Central Nervous SystemMusfira KhalidNo ratings yet
- Lecture 8 Special Senses Part 3-VisionDocument32 pagesLecture 8 Special Senses Part 3-VisionMoses Jr KazevuNo ratings yet
- Understanding the Cardiovascular SystemDocument7 pagesUnderstanding the Cardiovascular SystemKhrean Kae SantiagoNo ratings yet
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (378)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Inside of a Dog: What Dogs See, Smell, and KnowFrom EverandInside of a Dog: What Dogs See, Smell, and KnowRating: 4 out of 5 stars4/5 (390)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- Why We Sleep: Unlocking the Power of Sleep and DreamsFrom EverandWhy We Sleep: Unlocking the Power of Sleep and DreamsRating: 4.5 out of 5 stars4.5/5 (2083)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (4)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- The Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsFrom EverandThe Dog Who Couldn't Stop Loving: How Dogs Have Captured Our Hearts for Thousands of YearsNo ratings yet
- The Invention of Tomorrow: A Natural History of ForesightFrom EverandThe Invention of Tomorrow: A Natural History of ForesightRating: 4.5 out of 5 stars4.5/5 (5)