You might also like
- The Ordinary Products ConflictsDocument2 pagesThe Ordinary Products ConflictsUngureanu Raluca100% (1)
- 1 - Vitamins MCQDocument6 pages1 - Vitamins MCQSubash KumarNo ratings yet
- Chapter 4 - MicronutrientsDocument114 pagesChapter 4 - MicronutrientsSami InglesNo ratings yet
- Fat-Soluble Vitamins Assignment) EdittedDocument33 pagesFat-Soluble Vitamins Assignment) EdittedDianzerShengNo ratings yet
- BIOS LIFE - Cleveland Clinic Trial by Dr. Dennis SprecherDocument5 pagesBIOS LIFE - Cleveland Clinic Trial by Dr. Dennis SprecherHisWellnessNo ratings yet
- Kelsey M. Cochrane, Rajavel Elango, Angela M. Devlin, Chantal Mayer, Jennifer A. Hutcheon and Crystal D. KarakochukDocument11 pagesKelsey M. Cochrane, Rajavel Elango, Angela M. Devlin, Chantal Mayer, Jennifer A. Hutcheon and Crystal D. KarakochukMatheus SouzaNo ratings yet
- Validity of Food Frequency Questionnaire Estimated Intakes of Folate and Other B Vitamins in A Region Without Folic Acid FortificationDocument10 pagesValidity of Food Frequency Questionnaire Estimated Intakes of Folate and Other B Vitamins in A Region Without Folic Acid FortificationKuro ChanNo ratings yet
- Plasma Total Homocysteine Status of Vegetarians Compared With OmnivoresDocument10 pagesPlasma Total Homocysteine Status of Vegetarians Compared With Omnivoresnta.gloffkaNo ratings yet
- Vitamin B ComplexDocument13 pagesVitamin B ComplexDilshad TasnimNo ratings yet
- Epigenetic Mechanisms Elicited by Nutrition in Early LifeDocument8 pagesEpigenetic Mechanisms Elicited by Nutrition in Early Lifehou1212!No ratings yet
- Am J Clin Nutr 2012 Theytaz 1008 16Document9 pagesAm J Clin Nutr 2012 Theytaz 1008 16evaNo ratings yet
- Nutritional "Omics" Technologies For Elucidating The Role(s) of Bioactive Food Components in Colon Cancer PreventionDocument5 pagesNutritional "Omics" Technologies For Elucidating The Role(s) of Bioactive Food Components in Colon Cancer PreventionJosé Carlos Rosas CarhuanchoNo ratings yet
- UK Food Standards Agency Cis-Monounsaturated FattyDocument7 pagesUK Food Standards Agency Cis-Monounsaturated FattybicycloonNo ratings yet
- Bhat Et Al., 2009Document5 pagesBhat Et Al., 2009Caoimhe O'BrienNo ratings yet
- Antidiabetic Properties of Dietary Flavonoids: A Cellular Mechanism ReviewDocument20 pagesAntidiabetic Properties of Dietary Flavonoids: A Cellular Mechanism ReviewRasha Samir SryoNo ratings yet
- Sirtori2012 Efectos de La Proteína de Altramuz y Arveja en Individuos Con Hipertensión LeveDocument8 pagesSirtori2012 Efectos de La Proteína de Altramuz y Arveja en Individuos Con Hipertensión Leveconsuelo aguilarNo ratings yet
- Sunday O. Oyedemi - Godswill Nwaogu - Chika I. Chukwuma - Olaoluwa T. Adeyemi - Motlalepula G. Matsabisa - Shasank S. Swain - Olayinka A. AiyegoroDocument16 pagesSunday O. Oyedemi - Godswill Nwaogu - Chika I. Chukwuma - Olaoluwa T. Adeyemi - Motlalepula G. Matsabisa - Shasank S. Swain - Olayinka A. AiyegoroGiovanni OliveiraNo ratings yet
- Uses of Vitamin b6Document14 pagesUses of Vitamin b6ShannonNo ratings yet
- Choline Homocysteine and PregnancyDocument2 pagesCholine Homocysteine and PregnancyNa OliveiraNo ratings yet
- 2019 McNultyDocument14 pages2019 McNultybryaneatscamerasNo ratings yet
- Antocinaninas Reduzem Risco de DM2 Am J Clin Nutr-2012Document9 pagesAntocinaninas Reduzem Risco de DM2 Am J Clin Nutr-2012Elenice Peltz NutricionistaNo ratings yet
- Metabolites 12 00927 v3Document20 pagesMetabolites 12 00927 v3adanNo ratings yet
- Dieta Baja en CarbohidratosDocument19 pagesDieta Baja en CarbohidratosESTUARDO ENRIQUE JO ORELLANANo ratings yet
- MMC 8Document27 pagesMMC 8Neil Patrick AngelesNo ratings yet
- Accepted Manuscript: 10.1016/j.mam.2016.11.004Document17 pagesAccepted Manuscript: 10.1016/j.mam.2016.11.004Muhammad TahirNo ratings yet
- Is Folic Acid Good For Everyone?: CommentaryDocument17 pagesIs Folic Acid Good For Everyone?: CommentaryKirubakaranNo ratings yet
- Theresa Davis USDA Grant Through 3:2019Document5 pagesTheresa Davis USDA Grant Through 3:2019The Nutrition CoalitionNo ratings yet
- Farooqi & O'Rahilly, 2009 - Leptin, A Pivotal Regulator of Human Energy HomeostasisDocument5 pagesFarooqi & O'Rahilly, 2009 - Leptin, A Pivotal Regulator of Human Energy HomeostasisMara ParamoNo ratings yet
- Nutrient Requirements and Optimal NutritionDocument6 pagesNutrient Requirements and Optimal NutritionNovriantiHawariniNo ratings yet
- Referencia 6. McNulty 2019Document14 pagesReferencia 6. McNulty 2019Erie RegaladoNo ratings yet
- HipoglikemiaDocument8 pagesHipoglikemiawilmaNo ratings yet
- Associations Between Folate and Choline Intake, Homocysteine Metabolism, and Genetic Polymorphism of MTHFR, BHMT and PEMT in Healthy Pregnant Polish WomenDocument5 pagesAssociations Between Folate and Choline Intake, Homocysteine Metabolism, and Genetic Polymorphism of MTHFR, BHMT and PEMT in Healthy Pregnant Polish WomenMauro Porcel de PeraltaNo ratings yet
- Am J Clin Nutr 2011 Verhoef 1 2Document2 pagesAm J Clin Nutr 2011 Verhoef 1 2Sugiyanto GunadiNo ratings yet
- RPrinz2009 6s - 5-Methyltetrahydrofolate Increases PlasmaDocument8 pagesRPrinz2009 6s - 5-Methyltetrahydrofolate Increases PlasmaAna Muñoa DuchateauNo ratings yet
- Artical Vit b6Document18 pagesArtical Vit b6sajjad khanNo ratings yet
- 1 s2.0 S2161831322009838 MainDocument14 pages1 s2.0 S2161831322009838 MainReni Puspita SariNo ratings yet
- 10 1016@j Heliyon 2020 E03632Document13 pages10 1016@j Heliyon 2020 E03632Tesa RafkhaniNo ratings yet
- Long-Term Exposure of INS-1 Rat Insulinoma Cells To Linoleic Acid and Glucose in Vitro Affects Cell Viability and Function Through Mitochondrial-Mediated PathwaysDocument11 pagesLong-Term Exposure of INS-1 Rat Insulinoma Cells To Linoleic Acid and Glucose in Vitro Affects Cell Viability and Function Through Mitochondrial-Mediated Pathwaysbharata.ishaya8999No ratings yet
- Is Folic Acid Good For Everyone?: CommentaryDocument17 pagesIs Folic Acid Good For Everyone?: CommentaryNanda Rizky FathiyaNo ratings yet
- Folic Acid in Pregnancy: Ethics/EducationDocument3 pagesFolic Acid in Pregnancy: Ethics/EducationAjengNo ratings yet
- Diet Rich in High Glucoraphanin Broccoli Reduces Plasma LDL Cholesterol: Evidence From Randomised Controlled TrialsDocument9 pagesDiet Rich in High Glucoraphanin Broccoli Reduces Plasma LDL Cholesterol: Evidence From Randomised Controlled TrialsYuni SahoeNo ratings yet
- Saengkaew 2017Document8 pagesSaengkaew 2017Tanya MorarNo ratings yet
- Research ArticleDocument6 pagesResearch ArticleKorry Meliana PangaribuanNo ratings yet
- Artigo 10 - Nutrients-13-01244Document20 pagesArtigo 10 - Nutrients-13-01244Pedro AfonsoNo ratings yet
- 5th Amino Acid Assessment WorkshopDocument12 pages5th Amino Acid Assessment WorkshopLetícia PachecoNo ratings yet
- Vitamin K and Osteoporosis: Nutraceutical Profile (September 2009)Document12 pagesVitamin K and Osteoporosis: Nutraceutical Profile (September 2009)Dinda AprilianiNo ratings yet
- ARTICULO FINAL (Agos 18)Document11 pagesARTICULO FINAL (Agos 18)Jaan Miguel Piñon CruzNo ratings yet
- Effects of Dietary Carbohydrate Restriction With High Protein Intake On Protein Metabolism and The Somatotropic AxisDocument7 pagesEffects of Dietary Carbohydrate Restriction With High Protein Intake On Protein Metabolism and The Somatotropic Axisthe bungalowvideotapesNo ratings yet
- Cornelis 2016Document11 pagesCornelis 2016Bee GuyNo ratings yet
- Estudo Coenzimaq10 PDFDocument12 pagesEstudo Coenzimaq10 PDFDanielle AliceNo ratings yet
- 1532502121article pdf555035515Document14 pages1532502121article pdf555035515SPN CDYNo ratings yet
- Inter-Organ Cross-Talk in Metabolic Syndrome: Christina Priest and Peter TontonozDocument12 pagesInter-Organ Cross-Talk in Metabolic Syndrome: Christina Priest and Peter TontonozxjorgwxNo ratings yet
- Intestinal Microbial Metabolism of Phosphatidylcholine and Cardiovascular RiskDocument10 pagesIntestinal Microbial Metabolism of Phosphatidylcholine and Cardiovascular RiskLeonardoNo ratings yet
- Modulation of Metabolic Detoxification Pathways Using Foods and Food-Derived Components - A Scientific Review With Clinical ApplicationDocument24 pagesModulation of Metabolic Detoxification Pathways Using Foods and Food-Derived Components - A Scientific Review With Clinical ApplicationkdfqhcutoxtvrbmicaNo ratings yet
- Nutrients 13 03965 v2Document11 pagesNutrients 13 03965 v2VaniNo ratings yet
- Am J Clin Nutr 2014 Lecoultre 268 75Document8 pagesAm J Clin Nutr 2014 Lecoultre 268 75Kang Opik TaufikNo ratings yet
- FosfatidilcolinaDocument6 pagesFosfatidilcolinaArthur FernandoNo ratings yet
- Ruth 2013Document17 pagesRuth 2013Paul SimononNo ratings yet
- Gill 2018Document20 pagesGill 2018Khumaira SantaNo ratings yet
- Absorption, Bioavailability, and Metabolism of FlavonoidsDocument12 pagesAbsorption, Bioavailability, and Metabolism of FlavonoidsM DhivagarNo ratings yet
- Sun 2019 Metformin Triggers Pyy Secretion inDocument7 pagesSun 2019 Metformin Triggers Pyy Secretion inMuôngNo ratings yet
- The Role and Influence of Gut MicrobiotaDocument7 pagesThe Role and Influence of Gut MicrobiotaMai NabiehNo ratings yet
- Ketogenic Diets Enhance Oxidative Stress and RadioChemo-Therapy Responses in Lung Cancer XenograftsDocument9 pagesKetogenic Diets Enhance Oxidative Stress and RadioChemo-Therapy Responses in Lung Cancer XenograftsCata AjuaNo ratings yet
- Dietary Fibre Functionality in Food and Nutraceuticals: From Plant to GutFrom EverandDietary Fibre Functionality in Food and Nutraceuticals: From Plant to GutFarah HosseinianNo ratings yet
- Blackmores - Apotek My PharmaDocument12 pagesBlackmores - Apotek My PharmaApotek MyPharmaNo ratings yet
- Harga Ecataloge (00) SD Maret 2021 PalembangDocument2 pagesHarga Ecataloge (00) SD Maret 2021 PalembangFarmasi beltimNo ratings yet
- HakdoggggDocument2 pagesHakdoggggHarry Venzon JaboliNo ratings yet
- Full Download Book Advanced Nutrition and Human Metabolism 3 PDFDocument41 pagesFull Download Book Advanced Nutrition and Human Metabolism 3 PDFwilliam.pilgrim541100% (22)
- Vitamins For BSC Nursing Students (PHARMACOLOGY) - By: BINI P SAMUEL, ASSISTANT PROFESSORDocument17 pagesVitamins For BSC Nursing Students (PHARMACOLOGY) - By: BINI P SAMUEL, ASSISTANT PROFESSORbinipsamuel25No ratings yet
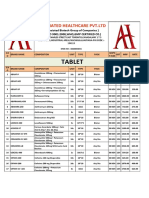
- Product List of Associated Healthcare - PCDDocument9 pagesProduct List of Associated Healthcare - PCDAshish SharmaNo ratings yet
- Ofertas de Farmacia 20% para AsociadosDocument2 pagesOfertas de Farmacia 20% para AsociadosCesar Caceres OrtelladoNo ratings yet
- Engleza Vitamina D Beneficii Avantaje Si DezavantajeDocument2 pagesEngleza Vitamina D Beneficii Avantaje Si DezavantajeLoredana LoreNo ratings yet
- BIOCHEM - ASSIGNMENT - Fernando, Kristel Cyrene DDocument9 pagesBIOCHEM - ASSIGNMENT - Fernando, Kristel Cyrene DKristel Cyrene FernandoNo ratings yet
- B-Complex Vitamins - Benefits, Side Effects and DosageDocument11 pagesB-Complex Vitamins - Benefits, Side Effects and DosageMohammed shamiul ShahidNo ratings yet
- RDA Chart RDA TableDocument4 pagesRDA Chart RDA TableVisshan SpirulinaNo ratings yet
- 33333determination of Water - and Fat-Soluble Vitamins in D555i ErentDocument21 pages33333determination of Water - and Fat-Soluble Vitamins in D555i ErentmohammaddammagNo ratings yet
- Contoh SURVEY GIZIDocument12 pagesContoh SURVEY GIZINandya AgustinaNo ratings yet
- Recommended Dietary Allowances - VitaminasDocument3 pagesRecommended Dietary Allowances - VitaminasDaniel Eduardo Lopez CabraNo ratings yet
- Dipa Escolab List ProductDocument4 pagesDipa Escolab List ProductRahmad HidayatNo ratings yet
- (C and D) of Nutrition AssessmentDocument44 pages(C and D) of Nutrition AssessmentNadia AbbasNo ratings yet
- TELUGU - Life Skill ManualDocument44 pagesTELUGU - Life Skill ManualChenna VijayNo ratings yet
- Pengaruh Kadar Glukosa Urine Metode Benedict, Fehling Dan Stick Setelah Ditambahkan Vitamin C DOSIS TINGGI/ 1000 MGDocument5 pagesPengaruh Kadar Glukosa Urine Metode Benedict, Fehling Dan Stick Setelah Ditambahkan Vitamin C DOSIS TINGGI/ 1000 MGRmf kumuay woffNo ratings yet
- NUTRITIONDocument26 pagesNUTRITIONJoeven HilarioNo ratings yet
- VITAMINS - StomDocument45 pagesVITAMINS - StomMihalachi-Anghel MariaNo ratings yet
- Bitot's SpotDocument19 pagesBitot's Spotyudhisthir panthiNo ratings yet
- The Vitamins Fifth Edition Fundamental Aspects in Nutrition and Health Gerald F Combs JR All ChapterDocument67 pagesThe Vitamins Fifth Edition Fundamental Aspects in Nutrition and Health Gerald F Combs JR All Chapteramanda.dangerfield238100% (6)
- Kinetic Study of Determination The Reaction Rate of Vitamin C in BrokoliDocument7 pagesKinetic Study of Determination The Reaction Rate of Vitamin C in BrokoliSyafa AsyharNo ratings yet
- Vitamin D Deficiency Is Associated With Increased Length of Stay After Acute Burn Injury: A Multicenter AnalysisDocument5 pagesVitamin D Deficiency Is Associated With Increased Length of Stay After Acute Burn Injury: A Multicenter Analysisdds (TMU)No ratings yet
- VitaminsDocument15 pagesVitaminsSAURABH PRAJAPATINo ratings yet
- ALbry t0 0 - DetailedDocument10 pagesALbry t0 0 - Detailedtopel manullangNo ratings yet