You might also like
- Enhancing Viability of Dried Lactic Acid Bacteria Prepared by Freeze Drying and Spray Drying Via Heat PreadaptationDocument9 pagesEnhancing Viability of Dried Lactic Acid Bacteria Prepared by Freeze Drying and Spray Drying Via Heat PreadaptationStephanie CaroNo ratings yet
- Mejor Malto A 4 °CDocument11 pagesMejor Malto A 4 °CLuisa TorresNo ratings yet
- Role of ProtectiveDocument8 pagesRole of ProtectiveAgus Prihanto PurnomoNo ratings yet
- LAB Lyophilization (PDF - Io)Document14 pagesLAB Lyophilization (PDF - Io)PHI LUUNo ratings yet
- Encapsulation of Lactobacillus Acidophilus in Moist-Heat-ResistantDocument8 pagesEncapsulation of Lactobacillus Acidophilus in Moist-Heat-ResistantMIRIAM CRISTINA DA SILVA GOMESNo ratings yet
- Effect of Cryopreservation Protocols On The Phenotypic Stability of YeastDocument7 pagesEffect of Cryopreservation Protocols On The Phenotypic Stability of Yeastandrea galibay perezNo ratings yet
- Postharvest Biology and TechnologyDocument8 pagesPostharvest Biology and TechnologyJoao Lecca RuízNo ratings yet
- Survival and Stability of Lactobacillus Fermentum and Wickerhamomyces Anomalus Strains Upon Lyophilisation With Different Cryoprotectant AgentsDocument5 pagesSurvival and Stability of Lactobacillus Fermentum and Wickerhamomyces Anomalus Strains Upon Lyophilisation With Different Cryoprotectant AgentsStephanie CaroNo ratings yet
- Cryobiology: Lactobacillus Fermentum L2Document9 pagesCryobiology: Lactobacillus Fermentum L2Stephanie CaroNo ratings yet
- Osmotic Dehydration Combined With Microwave Drying of Tilapia Fillets The Impacts of Osmosis DehydraDocument10 pagesOsmotic Dehydration Combined With Microwave Drying of Tilapia Fillets The Impacts of Osmosis Dehydrafrancisco cadenaNo ratings yet
- Food HydrocolloidsDocument9 pagesFood HydrocolloidsRSNo ratings yet
- 1 s2.0 S0958694616300772 Main PDFDocument8 pages1 s2.0 S0958694616300772 Main PDFhergofraNo ratings yet
- 10.1007@s11947 019 02340 9Document11 pages10.1007@s11947 019 02340 9Vy NguyễnNo ratings yet
- Jurnal Mikroba YogurtDocument7 pagesJurnal Mikroba YogurtsoniaaaniaaaNo ratings yet
- S7 HP Frozen Carrot Juice 2017Document7 pagesS7 HP Frozen Carrot Juice 2017MeLissa AymNo ratings yet
- Ahmad Et Al 2020 Temperature Dependent Morphological Changes On Algal GrowthDocument12 pagesAhmad Et Al 2020 Temperature Dependent Morphological Changes On Algal GrowthanaideNo ratings yet
- Shoji 2013Document6 pagesShoji 201315041168No ratings yet
- Journal of King Saud University - Science: Original ArticleDocument8 pagesJournal of King Saud University - Science: Original ArticleAri tiwiNo ratings yet
- Flow-Cytometric Assessment of Damages To Acetobacter Senegalensis During Freeze-Drying Process and StorageDocument10 pagesFlow-Cytometric Assessment of Damages To Acetobacter Senegalensis During Freeze-Drying Process and StorageJamesNo ratings yet
- Preservation of Probiotic Strains Isolated From Kefir by Spray DryingDocument6 pagesPreservation of Probiotic Strains Isolated From Kefir by Spray DryingItta RahmaliaNo ratings yet
- Smet 2016Document10 pagesSmet 2016Meme WindNo ratings yet
- Kultur Akar Adventif 2Document10 pagesKultur Akar Adventif 2PUSAT INFORMASINo ratings yet
- Evaluation of Proline, Soluble Sugar and ABA Content in SoybeanDocument10 pagesEvaluation of Proline, Soluble Sugar and ABA Content in SoybeanWildanul HasanahNo ratings yet
- Effects of Different Thawing Methods On Physicochemical Properties and Structure of Largemouth Bass (Micropterus Salmoides)Document10 pagesEffects of Different Thawing Methods On Physicochemical Properties and Structure of Largemouth Bass (Micropterus Salmoides)Juan Diego Espinoza AlvinesNo ratings yet
- Tu Et Al, 2015. Effects of Different Freezing Methods On The Quality and Microstructure of Lotus (Nelumbo Nucifera) RootDocument7 pagesTu Et Al, 2015. Effects of Different Freezing Methods On The Quality and Microstructure of Lotus (Nelumbo Nucifera) RootHardyantiPutriNo ratings yet
- Journal of The Saudi Society of Agricultural SciencesDocument7 pagesJournal of The Saudi Society of Agricultural SciencesFERNANDO PATARROYONo ratings yet
- Osmodehidratacion ManzanaDocument9 pagesOsmodehidratacion ManzanaPedro MedinaNo ratings yet
- Alvarez 2008Document8 pagesAlvarez 2008DarthNo ratings yet
- Effect of Sperm Storage and Selection Techniques On Sperm ParametersDocument13 pagesEffect of Sperm Storage and Selection Techniques On Sperm ParametersgrimmNo ratings yet
- Uji Kadar Air (Jurin)Document11 pagesUji Kadar Air (Jurin)HANIP GALIH PAMBUDINo ratings yet
- Stress Adaptation and Cross-Protection of Lactobacillus Plantarum KLDS 1.0628Document10 pagesStress Adaptation and Cross-Protection of Lactobacillus Plantarum KLDS 1.0628momo boulkNo ratings yet
- High Pressure Processing A Potential Technology For Processing and Preservation Diary FoodDocument10 pagesHigh Pressure Processing A Potential Technology For Processing and Preservation Diary FoodKim Anh HaNo ratings yet
- Characterization of Novel Probiotic Nanocomposite Films Based On NanoDocument11 pagesCharacterization of Novel Probiotic Nanocomposite Films Based On NanoFadil NurhidayatNo ratings yet
- DSC As A Tool To Assess Physiological Evolution of Apples PreservedDocument6 pagesDSC As A Tool To Assess Physiological Evolution of Apples PreservedElkyn BohórquezNo ratings yet
- Low-Moisture Food Matrices As Probiotic CarriersDocument11 pagesLow-Moisture Food Matrices As Probiotic CarriersgraceNo ratings yet
- JRPS 7 31Document6 pagesJRPS 7 31anggrek IPBNo ratings yet
- 1 s2.0 S0304423815001375 MainDocument5 pages1 s2.0 S0304423815001375 MainFrancisco Herrera DalletoNo ratings yet
- 8261 10.1016@j.jfoodeng.2019.03.011Document34 pages8261 10.1016@j.jfoodeng.2019.03.011Renaldi ArmanNo ratings yet
- Effect of Freezing Rates and Freeze-Thaw Cycles On The Texture, Microstructure and Pectic Substances of MangoDocument8 pagesEffect of Freezing Rates and Freeze-Thaw Cycles On The Texture, Microstructure and Pectic Substances of MangoIlham Munawar MansyurNo ratings yet
- Material 3744Document11 pagesMaterial 3744samy cajusolNo ratings yet
- Acidophilus ParticulaDocument6 pagesAcidophilus ParticulaMay MolinaNo ratings yet
- 1 s2.0 S0308814621005872 MainDocument10 pages1 s2.0 S0308814621005872 MainNelluri PujaNo ratings yet
- Enhancing the Freezing-Thawing Tolerance of Frozen Dough Using Ε-poly-L-lysine Treated Yeast 2020Document7 pagesEnhancing the Freezing-Thawing Tolerance of Frozen Dough Using Ε-poly-L-lysine Treated Yeast 2020Abo Ahmad AlsaidiNo ratings yet
- 0fcfd5093e28d328a8000000 PDFDocument7 pages0fcfd5093e28d328a8000000 PDFricky hutagalungNo ratings yet
- Scientia Horticulturae: SciencedirectDocument7 pagesScientia Horticulturae: SciencedirectThiago NettoNo ratings yet
- Potential Life Cycle Carbon Savings For Immersion FreezingDocument12 pagesPotential Life Cycle Carbon Savings For Immersion FreezingNataliaNo ratings yet
- Foods: Osmodehydrofreezing: An Integrated Process For Food Preservation During Frozen StorageDocument25 pagesFoods: Osmodehydrofreezing: An Integrated Process For Food Preservation During Frozen StorageNia PutriNo ratings yet
- 1 s2.0 S0260877415003301 MainDocument8 pages1 s2.0 S0260877415003301 MainRSNo ratings yet
- Book ChapterDocument22 pagesBook ChapterHimani Singh GaurNo ratings yet
- Alternative Postharvest Pre Treatment Strategies For Quality and M - 2021 - HeliDocument9 pagesAlternative Postharvest Pre Treatment Strategies For Quality and M - 2021 - Heliمصطفى الذاريNo ratings yet
- Eichlerov 2014 NO PDFDocument10 pagesEichlerov 2014 NO PDFGUIDO ERNESTO VILLOTA CALVACHINo ratings yet
- Li 2018Document10 pagesLi 2018Prakoso AdiNo ratings yet
- Journal Pre-Proof: LWT - Food Science and TechnologyDocument46 pagesJournal Pre-Proof: LWT - Food Science and TechnologyJulio MoralesNo ratings yet
- Jurnal Anpang FixsDocument9 pagesJurnal Anpang FixsNabilla EvaNo ratings yet
- Articulo Operaciones UnitariasDocument3 pagesArticulo Operaciones UnitariasJahana GarciaNo ratings yet
- Lavoisier 2019Document10 pagesLavoisier 2019Jose de PapadopoulosNo ratings yet
- 脉冲电场预处理改善冻干植物材料的微观结构和松脆性:以草莓为例Document8 pages脉冲电场预处理改善冻干植物材料的微观结构和松脆性:以草莓为例blk wtNo ratings yet
- Effects of Thermal ProcessingDocument13 pagesEffects of Thermal ProcessingDome SalcedoNo ratings yet
- Effect of Processing and Packaging Conditions On Quality of Refrigerated Potato StripsDocument7 pagesEffect of Processing and Packaging Conditions On Quality of Refrigerated Potato Stripsolotu_olaseindeNo ratings yet
- Sri Sauhard Lakhanpal Vs State of UPDocument6 pagesSri Sauhard Lakhanpal Vs State of UPShreya SinhaNo ratings yet
- Lecture#1: Plant Cell and Tissue CultureDocument12 pagesLecture#1: Plant Cell and Tissue CultureIbrahimAslamNo ratings yet
- Phylogenetic Trees Activity - Farizah AliDocument2 pagesPhylogenetic Trees Activity - Farizah AliFarizah AliNo ratings yet
- Alzheimers-Disease-Genetics-Fact-Sheet 0Document8 pagesAlzheimers-Disease-Genetics-Fact-Sheet 0api-285676076No ratings yet
- BT503 - Past Papers PDFDocument83 pagesBT503 - Past Papers PDFJawad100% (1)
- CauveryMK - CVDocument3 pagesCauveryMK - CVMichelle KnightNo ratings yet
- Study Strategies For BMB 401 or How To Get A Good Grade in Biochemistry?Document6 pagesStudy Strategies For BMB 401 or How To Get A Good Grade in Biochemistry?E12H12No ratings yet
- Student Exploration: Building DNADocument4 pagesStudent Exploration: Building DNAShiper 33No ratings yet
- NematodeschapterDocument9 pagesNematodeschapterArjelyn Mae D BuendiaNo ratings yet
- Pneumacult™-Ali: For Bronchial Epithelial CellsDocument2 pagesPneumacult™-Ali: For Bronchial Epithelial CellsDunkMeNo ratings yet
- Parasitology: The Science Which Studying The Relationship Between Different TwoDocument10 pagesParasitology: The Science Which Studying The Relationship Between Different TwoFatima Ali Abdel-RedaNo ratings yet
- Anti HIV Using Nanorobots: Dr. Ankita Joshi, Akash PardeshiDocument7 pagesAnti HIV Using Nanorobots: Dr. Ankita Joshi, Akash PardeshiAmal Rejan Amal RejanNo ratings yet
- Nutrition Answers FinalDocument9 pagesNutrition Answers FinalelizabethNo ratings yet
- Instant Download Ebook PDF Essentials of Biology 4th Edition by Sylvia Mader PDF ScribdDocument41 pagesInstant Download Ebook PDF Essentials of Biology 4th Edition by Sylvia Mader PDF Scribdjune.thompson131100% (46)
- CattleDocument66 pagesCattleAlbyziaNo ratings yet
- Intestinal and Luminal ProtozoaDocument27 pagesIntestinal and Luminal ProtozoatasyaghazzanniNo ratings yet
- 9700 Y16 Sy 1Document81 pages9700 Y16 Sy 1api-255467062No ratings yet
- Prescotts Microbiology 10th Edition Willey Solutions Manual 1Document4 pagesPrescotts Microbiology 10th Edition Willey Solutions Manual 1ruth98% (48)
- Immunological Reviews - 2009 - Sayegh - The Chicken B Cell Receptor Complex and Its Role in Avian B Cell DevelopmentDocument14 pagesImmunological Reviews - 2009 - Sayegh - The Chicken B Cell Receptor Complex and Its Role in Avian B Cell DevelopmenthannahNo ratings yet
- Science 3-9 DLP Second QuarterDocument12 pagesScience 3-9 DLP Second QuarterCozette AtendidoNo ratings yet
- Obesity 1. Basic Science of ObesityDocument10 pagesObesity 1. Basic Science of ObesitygraceNo ratings yet
- Clinical Genomics Laboratory: Test ResultDocument1 pageClinical Genomics Laboratory: Test ResultPeds Lim PagayatanNo ratings yet
- Sexual and Asexual Reproduction Detailed Lesson PlanDocument9 pagesSexual and Asexual Reproduction Detailed Lesson PlanChristianJamesCapiz96% (25)
- BIO-Resonance Diagnostic Systems OBERON - Flyer v2Document3 pagesBIO-Resonance Diagnostic Systems OBERON - Flyer v2OBERON-INTROSPECT-BIOSPECTNo ratings yet
- CV Aeman 2018 NewDocument4 pagesCV Aeman 2018 NewAeman KhalidNo ratings yet
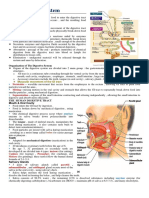
- The Digestive System MODULEDocument3 pagesThe Digestive System MODULEBen Moldenhaur100% (2)
- Biology Feynman TechniquesDocument9 pagesBiology Feynman TechniquesmuthulaxmiNo ratings yet
- Assessment: (Post-Test) : Directions: Read and Analyze Each Item Carefully. Select The Correct Answer From The ChoicesDocument3 pagesAssessment: (Post-Test) : Directions: Read and Analyze Each Item Carefully. Select The Correct Answer From The ChoicesChelleyOllitro100% (2)
- Compost and The Soil Food WebDocument7 pagesCompost and The Soil Food WebMoreMoseySpeed100% (2)
- Lecture D111L Week 05 S13Document64 pagesLecture D111L Week 05 S13Tahir RizviNo ratings yet