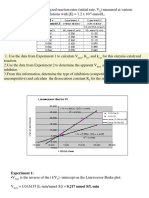
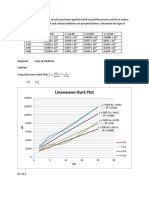
You might also like
- Chemical Equilibrium NotesDocument4 pagesChemical Equilibrium NotesHaile CordaNo ratings yet
- ch03 SM Chemistry2eDocument36 pagesch03 SM Chemistry2eLLL0% (1)
- Michaelis Menten EquationDocument9 pagesMichaelis Menten Equationsadaf zaidiNo ratings yet
- HGE Refresher QuestionsDocument8 pagesHGE Refresher QuestionsCj SuarezNo ratings yet
- IsomersDocument5 pagesIsomersSobia Saleem100% (1)
- My CalculationsDocument39 pagesMy CalculationsWaqas Khan ChannarNo ratings yet
- Chemical KineticsDocument169 pagesChemical KineticsHope WorldNo ratings yet
- Enzyme KineticsDocument13 pagesEnzyme KineticsalicjadzNo ratings yet
- DryerDocument80 pagesDryerAaron Paulo Bernardo100% (2)
- Hybridization 1Document37 pagesHybridization 1Anzari MuhammadNo ratings yet
- Problem Set - Enzymes From LehningerDocument11 pagesProblem Set - Enzymes From LehningervioletbrownNo ratings yet
- Amylase Assay 2Document9 pagesAmylase Assay 2Rahman ImudaNo ratings yet
- A.1. Competitive InhibitionDocument5 pagesA.1. Competitive InhibitionFlorecita Cabañog100% (1)
- Enzyme InhibitionDocument7 pagesEnzyme InhibitionJane Docdoc100% (1)
- Enzyme Kinetics ProblemsDocument5 pagesEnzyme Kinetics ProblemsBlessy Gabayno100% (1)
- Protein Structure and Folding LectureDocument18 pagesProtein Structure and Folding Lecturemaryscribd241No ratings yet
- A.2. Noncompetitive InhibitionDocument6 pagesA.2. Noncompetitive InhibitionFlorecita CabañogNo ratings yet
- V VI EnzymesDocument59 pagesV VI Enzymesthamizh555100% (2)
- 5 Enzyme Kinetics-InhibitionDocument40 pages5 Enzyme Kinetics-InhibitionJoel SmolanoffNo ratings yet
- Chapter 10 Chemical Kinetics IIDocument131 pagesChapter 10 Chemical Kinetics IIChicken ChickenNo ratings yet
- Lab ReportDocument8 pagesLab ReportUsman AliNo ratings yet
- MORE Practice For Michaelis-Menten ShitDocument3 pagesMORE Practice For Michaelis-Menten ShitMalcolm Charles100% (1)
- Chem 40 Enzyme KineticsDocument85 pagesChem 40 Enzyme KineticsJustine Grace Mariano100% (1)
- Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis IIDocument36 pagesSodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis IIYUEALVIN17No ratings yet
- 7-Bioc431 Enzymes Kinetics S15Document34 pages7-Bioc431 Enzymes Kinetics S15Shariq Mansoor KhanNo ratings yet
- Flash DesignDocument14 pagesFlash Designdavoin_88No ratings yet
- Chapter-0 Control Valves Handbook PDFDocument60 pagesChapter-0 Control Valves Handbook PDFOscarGomezMecanico100% (1)
- Assign#2 Enzyme KineticsDocument3 pagesAssign#2 Enzyme KineticsKaren Castro100% (2)
- Biochemistry II End of Semester Exam QuestionsDocument6 pagesBiochemistry II End of Semester Exam Questionsnima00No ratings yet
- Excel 2007 TutorialDocument5 pagesExcel 2007 TutorialromelcarvajalNo ratings yet
- Electronic Absorption Spectra and Geometry of Organic Molecules: An Application of Molecular Orbital TheoryFrom EverandElectronic Absorption Spectra and Geometry of Organic Molecules: An Application of Molecular Orbital TheoryRating: 5 out of 5 stars5/5 (1)
- Rotary Thermal Dryer Design Book - FeecoDocument9 pagesRotary Thermal Dryer Design Book - Feecorajeevup2004No ratings yet
- Name: Kristine Joy Atos Block: BSN 1-D Practice ProblemsDocument6 pagesName: Kristine Joy Atos Block: BSN 1-D Practice ProblemsJenz Hope Segui Novela100% (3)
- Acid Gas RemovalDocument18 pagesAcid Gas RemovalKailash NarainNo ratings yet
- Enzyme KineticsDocument3 pagesEnzyme KineticsZeny Naranjo100% (2)
- Introductory Course For Analytical ChemistryDocument3 pagesIntroductory Course For Analytical ChemistryJohn AguilaNo ratings yet
- 4.1.3 Properties of Coals PDFDocument7 pages4.1.3 Properties of Coals PDFJaco KotzeNo ratings yet
- Inorg Reactionn MechanismsDocument38 pagesInorg Reactionn MechanismsThabang MolekoNo ratings yet
- ENZYMES Problems and SolutionsDocument6 pagesENZYMES Problems and SolutionsChadby GraNaNo50% (2)
- Problem Set On Enzyme Kinetics - FS - 2012 - 2013Document2 pagesProblem Set On Enzyme Kinetics - FS - 2012 - 2013Chay Alcantara100% (1)
- Physical Properties:: Haldia RefineryDocument43 pagesPhysical Properties:: Haldia Refinerymujeebtalib100% (3)
- MIT Lecture 6 5.07 Biochemistry LectureDocument16 pagesMIT Lecture 6 5.07 Biochemistry LectureakiridoNo ratings yet
- Question CH06+answer PDFDocument8 pagesQuestion CH06+answer PDFCris-Anne Juangco III100% (1)
- Enzyme Kinetics Examples and ProblemsDocument4 pagesEnzyme Kinetics Examples and Problemskiiadizon07100% (1)
- Buffer SolutionDocument14 pagesBuffer SolutionSony0% (1)
- 3Document4 pages3biotech_vidhya100% (1)
- Functional AnFunctional Analysis Lecture Notes Jeff Schenkeralysis Jeff SchenkerDocument169 pagesFunctional AnFunctional Analysis Lecture Notes Jeff Schenkeralysis Jeff Schenkernim19870% (1)
- Voet - Chapt - 12 Properties of EnzymesDocument102 pagesVoet - Chapt - 12 Properties of Enzymestelmo flowNo ratings yet
- Stefan Boltzmann Law PDFDocument3 pagesStefan Boltzmann Law PDFESAKKIMALA SNo ratings yet
- Orgo Cheat Sheets 08 2019 PDFDocument34 pagesOrgo Cheat Sheets 08 2019 PDFKobe AcobNo ratings yet
- Boger CourseDocument477 pagesBoger CourseharrypoutreurNo ratings yet
- CH 8 - ProblemsDocument4 pagesCH 8 - ProblemsKhris GriffisNo ratings yet
- Enzyme KineticsDocument72 pagesEnzyme Kineticsitokki otoya100% (1)
- Unit 2 - Proteins NotesDocument16 pagesUnit 2 - Proteins NotesPRIYA SHEETAK100% (1)
- A2 4 1 1 Krebs CycleDocument12 pagesA2 4 1 1 Krebs CycleHo Man ChanNo ratings yet
- Thermochemistry: - Petrucci, Herring Madura and BissonnetteDocument49 pagesThermochemistry: - Petrucci, Herring Madura and BissonnetteYousif Khalid100% (1)
- Expts For Chem EnggDocument37 pagesExpts For Chem Enggblackbeauty140% (1)
- Structures of ProteinsDocument15 pagesStructures of ProteinsEdward Juanico HerradaNo ratings yet
- 14 - Diffusion Through A Spherical Stagnant FilmDocument16 pages14 - Diffusion Through A Spherical Stagnant FilmAvinnaash SureshNo ratings yet
- Alpha Carbon Chemistry - Enols and EnolatesDocument49 pagesAlpha Carbon Chemistry - Enols and EnolatesKuku MandavaNo ratings yet
- Kinetic Vs Chemical MechanismDocument34 pagesKinetic Vs Chemical MechanismIgnacio Bascuñán OyarceNo ratings yet
- Conjugated Dyes Lab EditedDocument8 pagesConjugated Dyes Lab EditedGugu Rutherford100% (1)
- Lecture 5a CatalysisDocument22 pagesLecture 5a CatalysisSandeep ChawlaNo ratings yet
- Metal Complexes or Coordination Compounds: Kfecn 4K Fe CNDocument90 pagesMetal Complexes or Coordination Compounds: Kfecn 4K Fe CNPavan Boro100% (1)
- Organic Chemistry II Chapter22Document8 pagesOrganic Chemistry II Chapter22RangikaNo ratings yet
- Exp5 Determination of Heat of Solution FDocument8 pagesExp5 Determination of Heat of Solution FJim100% (1)
- Octahedral Molecular GeometryDocument24 pagesOctahedral Molecular GeometryAnonymous gUjimJKNo ratings yet
- Medicinal Chemistry—III: Main Lectures Presented at the Third International Symposium on Medicinal ChemistryFrom EverandMedicinal Chemistry—III: Main Lectures Presented at the Third International Symposium on Medicinal ChemistryP. PratesiNo ratings yet
- (Lec) May 20Document32 pages(Lec) May 20Aldren RebaLdeNo ratings yet
- HysysDocument36 pagesHysysRamiro ArcentalesNo ratings yet
- 02 K Charts - AE+SIDocument4 pages02 K Charts - AE+SIMohd Muizzuddin WahabNo ratings yet
- Week 3 - Computer ActivityDocument2 pagesWeek 3 - Computer ActivityAldren RebaLdeNo ratings yet
- HxyDocument1 pageHxyAldren RebaLdeNo ratings yet
- Che 133 Submission Rubrics Ay1617Document2 pagesChe 133 Submission Rubrics Ay1617Aldren RebaLdeNo ratings yet
- Rev MTC For PackedDocument15 pagesRev MTC For PackedAldren RebaLdeNo ratings yet
- GEA Barr-Rosin - Industrial Drying Brochure - English - A4 - tcm11-16154Document12 pagesGEA Barr-Rosin - Industrial Drying Brochure - English - A4 - tcm11-16154Daniel SouzaNo ratings yet
- Ads or PtionDocument15 pagesAds or PtionAldren RebaLdeNo ratings yet
- Keyword: Thermal, Heat, Radiation, Infra-Red, EmitDocument24 pagesKeyword: Thermal, Heat, Radiation, Infra-Red, Emitvenkat_09No ratings yet
- Radiation Heat TransferDocument40 pagesRadiation Heat TransferAldren RebaLdeNo ratings yet
- BiochemDocument1 pageBiochemLiezyl Dela CruzNo ratings yet
- BSCE Curriculum Checklist 2012Document1 pageBSCE Curriculum Checklist 2012Rad Briones LibrandaNo ratings yet
- Lecture10 PDFDocument3 pagesLecture10 PDFAldren RebaLdeNo ratings yet
- Lecture Chapter 15Document48 pagesLecture Chapter 15Aldren RebaLdeNo ratings yet
- RDVFwallboard GypsumDocument14 pagesRDVFwallboard Gypsumkrishnabiju2010100% (1)
- Differential Pressure Flowmeters CHAP2Document1 pageDifferential Pressure Flowmeters CHAP2Aldren RebaLdeNo ratings yet
- XDocument16 pagesXAldren RebaLdeNo ratings yet
- Presented By: Jeanie Joy Mata Bs Che V School of Technology 15 August 2013Document30 pagesPresented By: Jeanie Joy Mata Bs Che V School of Technology 15 August 2013Aldren RebaLdeNo ratings yet
- Volumetric MeterDocument8 pagesVolumetric MeterAldren RebaLdeNo ratings yet
- Flue Gas Data (Final)Document4 pagesFlue Gas Data (Final)Aldren RebaLdeNo ratings yet
- Volumetric MeterDocument8 pagesVolumetric MeterAldren RebaLdeNo ratings yet
- 76575695Document9 pages76575695Aldren RebaLdeNo ratings yet
- BS Chem StudyPlanDocument2 pagesBS Chem StudyPlanAldren RebaLdeNo ratings yet
- Chap 014Document31 pagesChap 014Aldren RebaLdeNo ratings yet
- The Randomized Complete Block Design (R)Document18 pagesThe Randomized Complete Block Design (R)Aldren RebaLdeNo ratings yet
- Standard Operation ProceduresDocument2 pagesStandard Operation ProceduresMohd Yusman KhairilNo ratings yet
- Hedco Fired Heaters PDFDocument25 pagesHedco Fired Heaters PDFALONSO GOMEZNo ratings yet
- CFX Intro 13.0 L01 Intro CFDDocument16 pagesCFX Intro 13.0 L01 Intro CFDAiman LatifNo ratings yet
- Pollution ControlDocument48 pagesPollution ControlAndreas NeshNo ratings yet
- Question Bank-Fluid Power System EngineeringDocument3 pagesQuestion Bank-Fluid Power System EngineeringShashank ShastriNo ratings yet
- Lubrication and Defects in ExtrusionDocument4 pagesLubrication and Defects in ExtrusionmostafaNo ratings yet
- S01 SchematicDocument22 pagesS01 SchematicLuan MaxNo ratings yet
- Investigation Towards The Efficiency of A Multi-Cyclone Dust Separator in Biomass CombustionDocument59 pagesInvestigation Towards The Efficiency of A Multi-Cyclone Dust Separator in Biomass CombustionAyman RiyadhNo ratings yet
- Aust Domestic Copper Tube To AS1571Document2 pagesAust Domestic Copper Tube To AS1571starykltNo ratings yet
- Landscape Design: WC Credit 1 Points: 1-2Document10 pagesLandscape Design: WC Credit 1 Points: 1-2KareenaNo ratings yet
- ET412C Refrigeration Test RigDocument2 pagesET412C Refrigeration Test RigRizwan Shad0% (1)
- TablasDocument2 pagesTablasluisepigmeniobermejoNo ratings yet
- Gas Storage-Main PresentationDocument21 pagesGas Storage-Main PresentationMurad RustamliNo ratings yet
- J Cep 2004 05 009Document10 pagesJ Cep 2004 05 009Sahand NekoeianNo ratings yet
- A Case Study On Thermodynamic Analysis of Cogeneration Power Plant (IRJET-V2I9163)Document5 pagesA Case Study On Thermodynamic Analysis of Cogeneration Power Plant (IRJET-V2I9163)luis hyungNo ratings yet
- 23.05.2021 2020 Well-Wise Daily Geological Report For Drilling Wells (Status at 5:00 AM)Document1 page23.05.2021 2020 Well-Wise Daily Geological Report For Drilling Wells (Status at 5:00 AM)Manash HazarikaNo ratings yet
- Refrigerants: University of Khartoum Mechanical Engineering Dept. Refrigeration and Air ConditioningDocument31 pagesRefrigerants: University of Khartoum Mechanical Engineering Dept. Refrigeration and Air ConditioningPlötzlich TunNo ratings yet
- Hvac A2Document14 pagesHvac A2sultanhusssainNo ratings yet
- File-Sensus Model 243 RegulatorDocument12 pagesFile-Sensus Model 243 RegulatorNaNo RegentNo ratings yet
- Particulate Control-2 Fabric Filters Particulate Scrubbers: Prof. Dr. Dentel Notes and Prof. Dr. Chang-Yu WuDocument53 pagesParticulate Control-2 Fabric Filters Particulate Scrubbers: Prof. Dr. Dentel Notes and Prof. Dr. Chang-Yu WuTushar KadgaleNo ratings yet
- Engineering Aspects of Roller Drier, Spray Drier, Fluid Bed Drier and Tray DrierDocument19 pagesEngineering Aspects of Roller Drier, Spray Drier, Fluid Bed Drier and Tray DrierRonak RawatNo ratings yet
- PDFDocument4 pagesPDFPradeep KumarNo ratings yet
- Module 5. Corrosion Prevention and ControlDocument10 pagesModule 5. Corrosion Prevention and ControlJohn Lexmar LeynesNo ratings yet
- Technical Data Sheet (TDS) : Puma Premium Motor Oil 15W-40Document2 pagesTechnical Data Sheet (TDS) : Puma Premium Motor Oil 15W-40Mohamed IsmailNo ratings yet