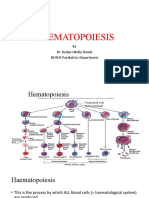
You might also like
- Ha Ema To PoiesisDocument38 pagesHa Ema To PoiesisEmma Joel OtaiNo ratings yet
- Normal HematopoesiseDocument35 pagesNormal HematopoesisesaketNo ratings yet
- Ch13 Hematopoiesis: Prepared and Presented byDocument47 pagesCh13 Hematopoiesis: Prepared and Presented bymmmmmmmmmmNo ratings yet
- Lym, Throm, GranuDocument12 pagesLym, Throm, Granumaxwell amponsahNo ratings yet
- Blood Formation LectureDocument24 pagesBlood Formation Lecturehassan aryaniNo ratings yet
- Lect 1Document47 pagesLect 1yamanassafNo ratings yet
- 1.anaemia Intro, Retic, Indices-1Document82 pages1.anaemia Intro, Retic, Indices-1InaGargNo ratings yet
- Bone Marrow and Hematopoiesis - 2Document7 pagesBone Marrow and Hematopoiesis - 2Sharan MurugaboopathyNo ratings yet
- Hematopoeitic System: Alya Amila Fitrie Lokot Donna LubisDocument80 pagesHematopoeitic System: Alya Amila Fitrie Lokot Donna LubisrubyniNo ratings yet
- Hematopoeitic System& Blood, KBK 2015 LDLDocument80 pagesHematopoeitic System& Blood, KBK 2015 LDLgita dwi ananda100% (1)
- Bs 260 HaematopoiesisDocument15 pagesBs 260 HaematopoiesisAkila AkinsNo ratings yet
- Hema Report PDFDocument25 pagesHema Report PDFdaliaNo ratings yet
- Spleen Functions & BonemarrowDocument4 pagesSpleen Functions & BonemarrowsnithajanNo ratings yet
- HEMA 1 Chapter 2Document4 pagesHEMA 1 Chapter 2GANASE, RONIA FENo ratings yet
- Exercise 5 HematopoiesisDocument11 pagesExercise 5 HematopoiesisAngela AninangNo ratings yet
- Week 1 - Study QuestionsDocument13 pagesWeek 1 - Study Questionsjess waldenNo ratings yet
- Hema TransesDocument25 pagesHema TransesNikoh Anthony EwayanNo ratings yet
- Bone MarrowDocument4 pagesBone MarrowSamra AwanNo ratings yet
- HEMA Lec 03 Bone Marrow Collection Lymphoid Organs 1Document9 pagesHEMA Lec 03 Bone Marrow Collection Lymphoid Organs 1Patrick AbellaNo ratings yet
- Bone Marrow: Andy Mclarnon, University of Birmingham, UkDocument2 pagesBone Marrow: Andy Mclarnon, University of Birmingham, Uknavita guptaNo ratings yet
- HEMATOPOIESIS StudentsDocument50 pagesHEMATOPOIESIS Studentskimberly abianNo ratings yet
- BSC Lecture Notes Semester 1Document7 pagesBSC Lecture Notes Semester 1satasha28No ratings yet
- Connective Tissues LectureDocument33 pagesConnective Tissues Lecturehassan aryaniNo ratings yet
- 1 Bone Tissue 2021-22Document35 pages1 Bone Tissue 2021-22Ali AfifNo ratings yet
- HematopoiesisDocument71 pagesHematopoiesisTalent AshjayNo ratings yet
- Anatomy and Physiology of Hemopoiesis The Bare Essentials: 1. Stem Cells Must ExistDocument4 pagesAnatomy and Physiology of Hemopoiesis The Bare Essentials: 1. Stem Cells Must ExistThomas Adi KhNo ratings yet
- Histologi Bone Marrow Feb 2019Document17 pagesHistologi Bone Marrow Feb 2019Listiana WindyNo ratings yet
- Hematology GNDocument94 pagesHematology GNJan Lian Enriquez NoolNo ratings yet
- Bio 2 Assignment 3Document16 pagesBio 2 Assignment 3Arun RajNo ratings yet
- HEMATOPOESISDocument71 pagesHEMATOPOESISHussenNo ratings yet
- Regulation of HematopoiesisDocument12 pagesRegulation of Hematopoiesisdeput_rprNo ratings yet
- Primary and Secondary Lymphoid Organs - Aditi SinghDocument50 pagesPrimary and Secondary Lymphoid Organs - Aditi SinghEunice PalloganNo ratings yet
- 2 - Introduction To Hematopoiesis and RBC ProductionDocument71 pages2 - Introduction To Hematopoiesis and RBC ProductionClaire GonoNo ratings yet
- Dev Skeletal SystemR2023Document83 pagesDev Skeletal SystemR2023amandagnaw19No ratings yet
- Seminar Submitted in Partial Fulfillment of Master Degree in Zoology OnDocument24 pagesSeminar Submitted in Partial Fulfillment of Master Degree in Zoology OnKhushbu BhardwajNo ratings yet
- Module Overview: Haematology and Transfusion Science (BMS 303)Document12 pagesModule Overview: Haematology and Transfusion Science (BMS 303)Grace PostmaNo ratings yet
- LECTURE 2 - HEMATOPOIESIS and ERYTHROPOIESIS - 10 - 17 - 2020Document49 pagesLECTURE 2 - HEMATOPOIESIS and ERYTHROPOIESIS - 10 - 17 - 2020apoorva krishnagiriNo ratings yet
- Jaringan Hemopoitik Darah: Disusun Oleh: Meidina Rahmawati (4401415078) Elvira Chrisma Aprilia F (4401415087)Document49 pagesJaringan Hemopoitik Darah: Disusun Oleh: Meidina Rahmawati (4401415078) Elvira Chrisma Aprilia F (4401415087)Suhaila Zakiya NajahNo ratings yet
- HEMATOPOIESISDocument7 pagesHEMATOPOIESISritaoktasariNo ratings yet
- Hematopoiesis 2022 VRDocument52 pagesHematopoiesis 2022 VRvictoria tavasNo ratings yet
- Chapter 20: The Cardiovascular System: Blood: Embryology Atlas HematopoiesisDocument8 pagesChapter 20: The Cardiovascular System: Blood: Embryology Atlas Hematopoiesisfjestefania6892No ratings yet
- The Hematologic SystemDocument7 pagesThe Hematologic SystemanyaNo ratings yet
- CLINPATH-01.Hematopoiesis & Morphology of Blood CellsDocument10 pagesCLINPATH-01.Hematopoiesis & Morphology of Blood CellsCharisse Angelica MacedaNo ratings yet
- Finals: Anatomy and Physiology Lecture: M4: L1. Hematopoeisis Hemopoietic Growth FactorsDocument33 pagesFinals: Anatomy and Physiology Lecture: M4: L1. Hematopoeisis Hemopoietic Growth FactorsEmily BernatNo ratings yet
- Genbio2 Week 2Document39 pagesGenbio2 Week 2Noel Vincent AgonoyNo ratings yet
- Haematology: Red Blood Cells DisordersDocument47 pagesHaematology: Red Blood Cells Disordersbv2328002No ratings yet
- Hematopoiesis: Pluri - Several) or Hemocytoblasts. These Cells Have The Capacity To Develop Into Many Different Types ofDocument12 pagesHematopoiesis: Pluri - Several) or Hemocytoblasts. These Cells Have The Capacity To Develop Into Many Different Types ofRajender ArutlaNo ratings yet
- Blood Lecture SlidesDocument144 pagesBlood Lecture Slidesgrace ncubeNo ratings yet
- Bone HistologyDocument50 pagesBone HistologyMajd HusseinNo ratings yet
- Embryo PresentationDocument12 pagesEmbryo PresentationLORENZO RAPHAEL ARAMILNo ratings yet
- Development of Blood Cells 2019Document31 pagesDevelopment of Blood Cells 2019Muhammad Anas Abbal100% (1)
- FALLSEM2023-24 BBIT305L TH VL2023240100183 2023-04-28 Reference-Material-IDocument9 pagesFALLSEM2023-24 BBIT305L TH VL2023240100183 2023-04-28 Reference-Material-IDrishti BisuiNo ratings yet
- 3 HEMA HematopoiesisDocument9 pages3 HEMA HematopoiesisJohanna Rose Cobacha SalvediaNo ratings yet
- Hematology Trans - Module 3 & 4 - Hematopoiesis and Lineage Specific HematopoiesisDocument6 pagesHematology Trans - Module 3 & 4 - Hematopoiesis and Lineage Specific HematopoiesisFaye Kyla Heart ResuelloNo ratings yet
- Lecture 28 NotesDocument6 pagesLecture 28 Notesapi-3721438100% (1)
- Blood and Lymph Nodes: P. Manyau School of Pharmacy University of ZimbabweDocument18 pagesBlood and Lymph Nodes: P. Manyau School of Pharmacy University of Zimbabwebrian mgabiNo ratings yet
- IUC - Haematology - Lecture 2 - Hemopoiesis-2021Document46 pagesIUC - Haematology - Lecture 2 - Hemopoiesis-2021Kingsly NdangaNo ratings yet
- Hematology Introduction: Organization of Blood and Blood Forming OrgansDocument50 pagesHematology Introduction: Organization of Blood and Blood Forming Organsmedun2009No ratings yet
- Red Blood Cells, Erthropoiesis&myeloid TissueDocument27 pagesRed Blood Cells, Erthropoiesis&myeloid TissueÉhmìđá LŷNo ratings yet
- Exercise 3 - Wireframe Geometry Creation and Editing - Rev ADocument33 pagesExercise 3 - Wireframe Geometry Creation and Editing - Rev AdevNo ratings yet
- Guru Stotram-1Document5 pagesGuru Stotram-1Green WattNo ratings yet
- Roy B. Kantuna, MPRM, CESE: (For Midterm) (Instructor)Document8 pagesRoy B. Kantuna, MPRM, CESE: (For Midterm) (Instructor)Athena Mae DitchonNo ratings yet
- John Deere CaseDocument2 pagesJohn Deere CaseAldo ReynaNo ratings yet
- Calydracomfort PiDocument16 pagesCalydracomfort PiionNo ratings yet
- Proceso Gmaw (RMD)Document8 pagesProceso Gmaw (RMD)Wilfredo RamirezNo ratings yet
- SIP Debugging Commands Overview - Cisco CommunityDocument5 pagesSIP Debugging Commands Overview - Cisco CommunitysenthilNo ratings yet
- Children Literature Evaluation Form I Aint Gonna Paint No MoreDocument4 pagesChildren Literature Evaluation Form I Aint Gonna Paint No Moreapi-548506674No ratings yet
- Portal 2 PresDocument15 pagesPortal 2 Presapi-666608332No ratings yet
- ECN 331 Fall 2018 SyllabusDocument6 pagesECN 331 Fall 2018 SyllabusDat BoiNo ratings yet
- Final Year Project Edi IrawanDocument75 pagesFinal Year Project Edi IrawanEdi IrawanNo ratings yet
- Henry's Bench: Keyes Ky-040 Arduino Rotary Encoder User ManualDocument4 pagesHenry's Bench: Keyes Ky-040 Arduino Rotary Encoder User ManualIsrael ZavalaNo ratings yet
- Sapper PamphletDocument24 pagesSapper Pamphletlonewolf2017100% (2)
- Miri Datalogger FlyerDocument1 pageMiri Datalogger FlyerernestoveigaNo ratings yet
- PlayStation MagazineDocument116 pagesPlayStation MagazineFrank Costello67% (3)
- Water Cement RatioDocument5 pagesWater Cement RatioCastro FarfansNo ratings yet
- HLSS 310 Critical Infrastructure ProtectionDocument12 pagesHLSS 310 Critical Infrastructure ProtectionMoffat HarounNo ratings yet
- The Macquarie Australian Slang DictionarDocument7 pagesThe Macquarie Australian Slang DictionarnetshidoNo ratings yet
- Leave Management System: Software Requirements Specification DocumentDocument6 pagesLeave Management System: Software Requirements Specification Documentk767No ratings yet
- TP PortalDocument25 pagesTP PortalSurendranNo ratings yet
- Test Bank For Organizations Behavior Structure Processes 14th Edition James GibsonDocument36 pagesTest Bank For Organizations Behavior Structure Processes 14th Edition James Gibsonfurrygrindlett3t1bt100% (37)
- Chapter 2 The Development of Evolutionary TheoryDocument5 pagesChapter 2 The Development of Evolutionary TheoryjohnNo ratings yet
- Letter of IntentDocument2 pagesLetter of Intentmansoorstanikzai4No ratings yet
- eLearnMarkets OptionsBuying HindiDocument17 pageseLearnMarkets OptionsBuying Hindisrinivas20% (1)
- Resume Arun.A: Career ObjectiveDocument2 pagesResume Arun.A: Career ObjectiveajithkumarNo ratings yet
- Computational/Experimental Analysis of Hydrokinetic TurbineDocument18 pagesComputational/Experimental Analysis of Hydrokinetic TurbineFozanNo ratings yet
- A Narrative Comprehensive Report of Student Teaching ExperiencesDocument82 pagesA Narrative Comprehensive Report of Student Teaching ExperiencesEpal Carlo74% (23)
- Cement Grouted Rock BoltsDocument28 pagesCement Grouted Rock BoltsBhaskar ReddyNo ratings yet
- ACS11Document3 pagesACS11Ebby OnyekweNo ratings yet
- Q1 WK 2 To 3 Las Fabm2 Kate DionisioDocument8 pagesQ1 WK 2 To 3 Las Fabm2 Kate DionisioFunji BuhatNo ratings yet