You might also like
- Canales IonicosDocument47 pagesCanales IonicosYvon Becerra CuevaNo ratings yet
- Ion Channel Factsbook: Extracellular Ligand-Gated ChannelsFrom EverandIon Channel Factsbook: Extracellular Ligand-Gated ChannelsNo ratings yet
- NT Release PPCNS 2023Document16 pagesNT Release PPCNS 2023Yasmin Sophia MNo ratings yet
- Shuai 2020Document5 pagesShuai 2020Virat KohNo ratings yet
- Lesson 4 Thyristors and TriacsDocument39 pagesLesson 4 Thyristors and TriacsChacko Mathew100% (1)
- B1 - 02 YorinakaDocument60 pagesB1 - 02 YorinakaWilan KrisnaNo ratings yet
- ELV1-L50-d Block-21092021 - 220419 - 143525Document12 pagesELV1-L50-d Block-21092021 - 220419 - 143525Aditya VijayvargiyaNo ratings yet
- ECE606: Solid State Devices: Bipolar Transistors A) Introduction B) Design (I)Document29 pagesECE606: Solid State Devices: Bipolar Transistors A) Introduction B) Design (I)Isaac BismonteNo ratings yet
- Lesson 5 Gate Turn Off Thyristor (GTO)Document23 pagesLesson 5 Gate Turn Off Thyristor (GTO)Chacko Mathew100% (1)
- 2 KineticsDocument18 pages2 KineticsLatifah FauziahNo ratings yet
- Lecture Outline03 ZhuDocument4 pagesLecture Outline03 Zhuapi-3700537No ratings yet
- 01-02 MS Ion Channels PDFDocument30 pages01-02 MS Ion Channels PDF16_dev5038No ratings yet
- 01-02 MS Ion Channels PDFDocument30 pages01-02 MS Ion Channels PDFSwaranjeet SinghNo ratings yet
- Technological Advances in Synthetic Biology: Subhayu BasuDocument33 pagesTechnological Advances in Synthetic Biology: Subhayu Basuapi-26720455No ratings yet
- PICASSO SNOLAB 12a 0Document44 pagesPICASSO SNOLAB 12a 0Ivan FelisNo ratings yet
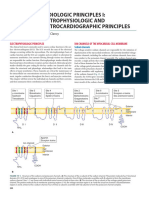
- Toxicología Goldfrank Capítulo de EKG en Paciente IntoxicadoDocument15 pagesToxicología Goldfrank Capítulo de EKG en Paciente IntoxicadoDavid Andrés BravoNo ratings yet
- Communication Systems, 5e: Chapter 11: Baseband Digital Transmission A. Bruce Carlson Paul B. CrillyDocument48 pagesCommunication Systems, 5e: Chapter 11: Baseband Digital Transmission A. Bruce Carlson Paul B. CrillySara JEBRINo ratings yet
- 3.8 Photosystem I Reduces NADP: StromaDocument14 pages3.8 Photosystem I Reduces NADP: StromaJack DanielNo ratings yet
- B5W4 Renal Regulation of Potassium Homeostasis STUDENTDocument29 pagesB5W4 Renal Regulation of Potassium Homeostasis STUDENTInhoNo ratings yet
- Principles of Semiconductor Devices-L27Document24 pagesPrinciples of Semiconductor Devices-L27LIAKMANNo ratings yet
- 3) Cell, Muscle, and Nerve PhysiologyDocument11 pages3) Cell, Muscle, and Nerve PhysiologyDION ANN SAYSONNo ratings yet
- BIOC441 Week 8 Slides Julia JooDocument17 pagesBIOC441 Week 8 Slides Julia JooJulia JooNo ratings yet
- Hydrocarbons HWDocument7 pagesHydrocarbons HWGagan PhadkeNo ratings yet
- 0 Introduction LTDocument34 pages0 Introduction LTrodrigoNo ratings yet
- Hydrocarbons HWDocument7 pagesHydrocarbons HWGagan PhadkeNo ratings yet
- Sodium: Segment Proximal Tubule Descending Limb Ascending Limb Distal Tubule Cortical CD Medullary CDDocument8 pagesSodium: Segment Proximal Tubule Descending Limb Ascending Limb Distal Tubule Cortical CD Medullary CDLoisirc M GoNo ratings yet
- Porphyry Deposit Alteration and Mineralization: Adi MaryonoDocument22 pagesPorphyry Deposit Alteration and Mineralization: Adi Maryonoirdan syafaatNo ratings yet
- Caso de Aplicación Dominios de Estimación Mineral Resource Estimation Springer Netherlands 2014Document12 pagesCaso de Aplicación Dominios de Estimación Mineral Resource Estimation Springer Netherlands 2014jorgeNo ratings yet
- Lecture Outline05 ZhuDocument4 pagesLecture Outline05 Zhuapi-3700537100% (2)
- Neural Signaling: The Membrane Potential: Lesson 9Document30 pagesNeural Signaling: The Membrane Potential: Lesson 9saavanakumarNo ratings yet
- SCR Diac TriacDocument7 pagesSCR Diac TriacSubaskaran SalavanNo ratings yet
- CHAPTER 7: DielectricsDocument23 pagesCHAPTER 7: DielectricsMoamen MohamedNo ratings yet
- Ion Channel Structure and FunctionDocument24 pagesIon Channel Structure and Functionshivanibhat1612No ratings yet
- Book References: Important Slides: ! Essential Cell BiologyDocument33 pagesBook References: Important Slides: ! Essential Cell BiologyМөнхгэрэл ГанбатNo ratings yet
- An Introduction To IGBT Operation: Application NoteDocument5 pagesAn Introduction To IGBT Operation: Application Noterodruren01No ratings yet
- Isomer & Reaksi OrganikDocument1 pageIsomer & Reaksi OrganiktekadputriwNo ratings yet
- NA08 DNA SequencingDocument3 pagesNA08 DNA SequencingAsmaa KhaledNo ratings yet
- Carbon Nanotube Field-Effect Transistors and Their Possible ApplicationsDocument36 pagesCarbon Nanotube Field-Effect Transistors and Their Possible ApplicationsAkshay PatharkarNo ratings yet
- Second LectrueDocument44 pagesSecond Lectrueapi-3700537No ratings yet
- TDC2h: Channels & PumpsDocument43 pagesTDC2h: Channels & Pumpsazazel666No ratings yet
- 2018 Review IDocument6 pages2018 Review IsophNo ratings yet
- 1 SPDocument43 pages1 SPMeghesh SamadhiyaNo ratings yet
- 2017 - EPS - Ravi Et AlDocument4 pages2017 - EPS - Ravi Et AlRavi GuptaNo ratings yet
- Thyristors and Triacs: DR - Arkan A.Hussein Power Electronics Fourth ClassDocument33 pagesThyristors and Triacs: DR - Arkan A.Hussein Power Electronics Fourth Classmohammed aliNo ratings yet
- Chanels+and+Transporters 2019Document58 pagesChanels+and+Transporters 201942030570No ratings yet
- Log BlankoDocument285 pagesLog BlankoFrengky BanunaekNo ratings yet
- Nervous SystemDocument49 pagesNervous Systemchudasamah680No ratings yet
- Solid State Chem PCHM 410Document57 pagesSolid State Chem PCHM 410Siva Kumar BellirajNo ratings yet
- Basic Concepts and Case Studies: High-Resolution X-Ray Scattering SpectrosDocument16 pagesBasic Concepts and Case Studies: High-Resolution X-Ray Scattering Spectrosuni137No ratings yet
- L-5 (DK) (Pe) ( (Ee) Nptel) 1Document5 pagesL-5 (DK) (Pe) ( (Ee) Nptel) 1GagneNo ratings yet
- ReceptorsDocument9 pagesReceptorsAlexandra AlexaNo ratings yet
- 7 - Carbene Carbyne Carbide ComplexesDocument8 pages7 - Carbene Carbyne Carbide ComplexesNandini GattadahalliNo ratings yet
- Brooks - Renal - Boards Step 1 ReviewDocument43 pagesBrooks - Renal - Boards Step 1 ReviewNyatarNo ratings yet
- Intro To Electronics P2 PDFDocument72 pagesIntro To Electronics P2 PDFSayan MandalNo ratings yet
- Supra Super Molecular Architecture DCDocument16 pagesSupra Super Molecular Architecture DCRaja LakshmiNo ratings yet
- How Cells WorkDocument27 pagesHow Cells WorkBensonNo ratings yet
- CHAPTER 6: DielectricsDocument36 pagesCHAPTER 6: DielectricsIstiaque AhmedNo ratings yet
- Complete Inorganic MarathonDocument407 pagesComplete Inorganic MarathonAdithya kumar JhaNo ratings yet
- Science Grade 9: Learning Activity SheetDocument6 pagesScience Grade 9: Learning Activity SheetChristian Ace Dequito RomeroNo ratings yet
- Brief Psychotic DisorderDocument3 pagesBrief Psychotic DisorderEugenio CainghugNo ratings yet
- Electromyography (EMG)Document6 pagesElectromyography (EMG)Tol Man ShresthaNo ratings yet
- Braztesol 2016 No Child Left Bored Iole VittiDocument43 pagesBraztesol 2016 No Child Left Bored Iole VittiGianna Saes Trevizani Negrinho GiannaNo ratings yet
- Health PsychologyDocument10 pagesHealth PsychologyDragana ModestyNo ratings yet
- Sem1 - Topic 9 - Geriatric Psychiatry-1Document36 pagesSem1 - Topic 9 - Geriatric Psychiatry-1prashant singhNo ratings yet
- Consuelo Gallegos-Ayala 2 1Document3 pagesConsuelo Gallegos-Ayala 2 1api-519799744No ratings yet
- 7 Habits Summary SheetDocument1 page7 Habits Summary Sheetjihardist100% (1)
- Explaining ADHD To TeachersDocument1 pageExplaining ADHD To TeachersChris100% (2)
- Instructional Design-Learning TheoryDocument32 pagesInstructional Design-Learning Theorymkderwish0% (1)
- Allie Cannon Anxiety Case StudyDocument9 pagesAllie Cannon Anxiety Case Studyapi-349885103No ratings yet
- Filipino Consumer Culture - DimalibotDocument4 pagesFilipino Consumer Culture - DimalibotKlenn Andrea Dimalibot100% (1)
- Language FlashcardsDocument52 pagesLanguage FlashcardsLoraNo ratings yet
- 4cog1 041150794 Tma1Document13 pages4cog1 041150794 Tma1Rajalingam RamanNo ratings yet
- Leadership PPT Chapter 2Document37 pagesLeadership PPT Chapter 2Arefeaine NegashNo ratings yet
- Quiz-08 25 23Document9 pagesQuiz-08 25 23Alexie AlmohallasNo ratings yet
- Does A Presentation's Medium Affect Its MessageDocument43 pagesDoes A Presentation's Medium Affect Its Messageellysa graceNo ratings yet
- Sample of Face Validity SummaryDocument3 pagesSample of Face Validity SummaryJohann Emmanuel MolatoNo ratings yet
- Emotional Regulation - RRLDocument3 pagesEmotional Regulation - RRLNaynay PaypayNo ratings yet
- Thinking About My Decision-MakingDocument18 pagesThinking About My Decision-MakingkrishnaNo ratings yet
- Communication Theories Perspectives Processes and Contexts PDFDocument2 pagesCommunication Theories Perspectives Processes and Contexts PDFBeverly43% (7)
- 15 Learning Theories in Education A Complete Summary PDFDocument24 pages15 Learning Theories in Education A Complete Summary PDFSilindelo NkosiNo ratings yet
- Physiology of VisionDocument49 pagesPhysiology of VisionPrakash PanthiNo ratings yet
- Ei PresentationDocument43 pagesEi PresentationRegine BulusanNo ratings yet
- Department of Education: Republic of The PhilippinesDocument3 pagesDepartment of Education: Republic of The PhilippinesGuia Marie Diaz BriginoNo ratings yet
- Scheff, T - Social Components in Depression, (2001) 64 Psychiatry 212Document13 pagesScheff, T - Social Components in Depression, (2001) 64 Psychiatry 212Sonja OsmanovićNo ratings yet
- 7 2 VoluntarismDocument31 pages7 2 Voluntarismuitsig.photoclubNo ratings yet
- Chapter 1: Introduction To Motivation and EmotionDocument9 pagesChapter 1: Introduction To Motivation and EmotionElla Rene MartinNo ratings yet
- Psychology - ESSENTIALS - Christian Bryan and Alan Law - Pearson 2013Document257 pagesPsychology - ESSENTIALS - Christian Bryan and Alan Law - Pearson 2013ibexamsNo ratings yet
- Conversion Disorder (Hysteria) : Functional Neurological Symptom DisorderDocument12 pagesConversion Disorder (Hysteria) : Functional Neurological Symptom DisorderSaman KhanNo ratings yet
- Lesson 3: The Human Person As An Embodied SpiritDocument2 pagesLesson 3: The Human Person As An Embodied SpiritSheryl NatinoNo ratings yet