You might also like
- 9 GlycogenDocument37 pages9 GlycogenShendi SuryanaNo ratings yet
- Chapter 16 Glycogen Metabolism and GluconeogenesisDocument32 pagesChapter 16 Glycogen Metabolism and GluconeogenesisSandraBadrousNo ratings yet
- Molarity, Molality, Normality, and Mass Percent Worksheet II Answer Key 11-12 PDFDocument3 pagesMolarity, Molality, Normality, and Mass Percent Worksheet II Answer Key 11-12 PDFGerald KamulanjeNo ratings yet
- Isolation of GlycogenDocument6 pagesIsolation of GlycogenAlleia Mae Urbano Mazo0% (2)
- Lesson 3 Glycogen-HZDocument20 pagesLesson 3 Glycogen-HZliaorNo ratings yet
- Glycogen Metabolism-FBADocument104 pagesGlycogen Metabolism-FBAozerbilge24No ratings yet
- GlycogenesisDocument7 pagesGlycogenesishammad646No ratings yet
- 8glycogen Metabolism2013 PDFDocument90 pages8glycogen Metabolism2013 PDFNazeer Baig MogalmearjaNo ratings yet
- Glycogen MetabolismDocument28 pagesGlycogen MetabolismshizarahimNo ratings yet
- 10-Glycogen MetabolismDocument23 pages10-Glycogen Metabolismshan.botan2003No ratings yet
- 2021 CHO Metabolism 2Document32 pages2021 CHO Metabolism 2xb2k9gzkc9No ratings yet
- GLIKOGENESISDocument45 pagesGLIKOGENESISChrista24796No ratings yet
- Glycogen, Glycogenolysis and GlycogenesisDocument8 pagesGlycogen, Glycogenolysis and GlycogenesisSierra OrtizNo ratings yet
- Glycogen MetabolismDocument34 pagesGlycogen MetabolismJomi PorinchuNo ratings yet
- Glycogen Metabolism: Unit Ii: Intermediary MetabolismDocument48 pagesGlycogen Metabolism: Unit Ii: Intermediary MetabolismSabha RabayaNo ratings yet
- Citric Acid Cycle: CharacteristicsDocument10 pagesCitric Acid Cycle: CharacteristicsYahya Daham Zafeer SakhrNo ratings yet
- Glycogenesis and GlycogenolysisDocument55 pagesGlycogenesis and Glycogenolysisclear mindNo ratings yet
- BiochemistryDocument9 pagesBiochemistryasish MohapatraNo ratings yet
- Carbohydrate MetabolismDocument57 pagesCarbohydrate MetabolismYulliza Kurniawaty L100% (1)
- (Bio3lec5) GlycogenDocument35 pages(Bio3lec5) GlycogenHerpy OtterNo ratings yet
- Glycogen MetabolismDocument27 pagesGlycogen Metabolismdrbishalchowdhury13No ratings yet
- Glycogen MetabolismDocument16 pagesGlycogen MetabolismMuhammad HashirNo ratings yet
- Presentation On Glycogen BreakdownDocument19 pagesPresentation On Glycogen BreakdownOmotayo oluwanifemiNo ratings yet
- Glycogen MetabolismDocument19 pagesGlycogen MetabolismEdward WinchesterrNo ratings yet
- Biochemistry Glycogen MetabolismDocument5 pagesBiochemistry Glycogen MetabolismPrashant Kumar SinghNo ratings yet
- Glycogen MetabolismDocument11 pagesGlycogen MetabolismSimra ZahidNo ratings yet
- Glycogen MetabolismDocument27 pagesGlycogen MetabolismDI NANo ratings yet
- Glycogen MetabolismDocument23 pagesGlycogen MetabolismAhsan Arshad BSIT-F16-LC-008No ratings yet
- LECTURE 3 - Carbohydrate MetabolismDocument40 pagesLECTURE 3 - Carbohydrate Metabolismmuhammedgmdidra95No ratings yet
- Important The 2 Mid Term Exam Is On November17 (Monday), 2014 in Remsen 017 at 5.00 PMDocument34 pagesImportant The 2 Mid Term Exam Is On November17 (Monday), 2014 in Remsen 017 at 5.00 PMjessjayleeNo ratings yet
- Met of CHODocument43 pagesMet of CHOKiya AlemuNo ratings yet
- 09-Biosynthesis of Carbohaydrates-2Document21 pages09-Biosynthesis of Carbohaydrates-2edh labNo ratings yet
- Carbohydrate Metabolism and BiosynthesisDocument44 pagesCarbohydrate Metabolism and BiosynthesisEmman RegondolaNo ratings yet
- 2.2 Carbohydrate Metabolism Part 2Document13 pages2.2 Carbohydrate Metabolism Part 2John Louis PulidoNo ratings yet
- Glycogen Metabolism.Document49 pagesGlycogen Metabolism.Santino MajokNo ratings yet
- Biosynthetic Pathways: Chemistry 203Document50 pagesBiosynthetic Pathways: Chemistry 203Jake OteroNo ratings yet
- Carbohydrate Metabolism (Unit 2) : Gluconeogenesis Continued .Document22 pagesCarbohydrate Metabolism (Unit 2) : Gluconeogenesis Continued .qwerty123No ratings yet
- Glycogenesis: DR Nazakat HussainDocument16 pagesGlycogenesis: DR Nazakat HussainMuhammad Jamal100% (1)
- Biochem TopicsDocument28 pagesBiochem TopicsLoraine ReuyanNo ratings yet
- Metabolisme GlikogenDocument17 pagesMetabolisme GlikogenMUHAMMAD RIDONo ratings yet
- Unit Ii: Bioenergetics and Carbohydrate MetabolismDocument51 pagesUnit Ii: Bioenergetics and Carbohydrate MetabolismErjel J. MalabananNo ratings yet
- Gluconeogenesis and GlycogenolysisDocument14 pagesGluconeogenesis and Glycogenolysissima mhammedNo ratings yet
- Metabolic Pathways For Carbohydrates: 7 Glycogen Metabolism 8 Gluconeogenesis: Glucose SynthesisDocument25 pagesMetabolic Pathways For Carbohydrates: 7 Glycogen Metabolism 8 Gluconeogenesis: Glucose SynthesisLa Ode RinaldiNo ratings yet
- Sinteza Si Depozitarea Carbohdratilor in Muschi Si FicatDocument17 pagesSinteza Si Depozitarea Carbohdratilor in Muschi Si FicatAnonymous UnVea6TnNo ratings yet
- GlycogenDocument25 pagesGlycogencandysunNo ratings yet
- Glycogen MetabolismDocument27 pagesGlycogen MetabolismRacheal MphandeNo ratings yet
- HB09 Glycogen MetabolismDocument59 pagesHB09 Glycogen MetabolismThisha MohanNo ratings yet
- Glycogenolysis and GlycogenesisDocument11 pagesGlycogenolysis and Glycogenesismiguel cuevas100% (2)
- Week 3 NotesDocument27 pagesWeek 3 NotesBae SeujiNo ratings yet
- Gandham. RajeevDocument14 pagesGandham. RajeevVishesh DwivediNo ratings yet
- 2020 Glycogen MetabolismDocument29 pages2020 Glycogen MetabolismChayanika KumarNo ratings yet
- Chapter 18 HandoutDocument8 pagesChapter 18 HandoutreynandNo ratings yet
- Assay of Tissue GlycogenDocument4 pagesAssay of Tissue GlycogentktchineeNo ratings yet
- 6 Glycogen 2017 MD PDFDocument36 pages6 Glycogen 2017 MD PDFIbrahim BarhamNo ratings yet
- L10 Glycogen and GluconeogenesisDocument25 pagesL10 Glycogen and GluconeogenesisCheng FuNo ratings yet
- 2.6 (BIOCHEMISTRY) Glycogen MetabolismDocument10 pages2.6 (BIOCHEMISTRY) Glycogen Metabolismlovelots1234No ratings yet
- Definition:: Fermentation)Document8 pagesDefinition:: Fermentation)Sreejith KrNo ratings yet
- Glycogen MetabolismDocument21 pagesGlycogen MetabolismKundaNo ratings yet
- GlycolysisDocument62 pagesGlycolysisafuaserwaa489No ratings yet
- MotivationDocument4 pagesMotivationMd. Muhaiminul IslamNo ratings yet
- Report FINAL CORRECTIONDocument44 pagesReport FINAL CORRECTIONMd. Muhaiminul IslamNo ratings yet
- URBS AssignmentDocument1 pageURBS AssignmentMd. Muhaiminul IslamNo ratings yet
- Topic 2, Control and Prevention of Microbial GrowthDocument23 pagesTopic 2, Control and Prevention of Microbial GrowthMd. Muhaiminul IslamNo ratings yet
- Undergraduate Thesis Proposal Form: M.A. Mukit 2.94 01750349993 Mohammad. Mohaimenul Islam 3.061 01671475244Document4 pagesUndergraduate Thesis Proposal Form: M.A. Mukit 2.94 01750349993 Mohammad. Mohaimenul Islam 3.061 01671475244Md. Muhaiminul IslamNo ratings yet
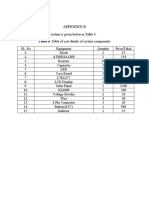
- Appendix B: Table 4: Table of Cost Details of System ComponentsDocument1 pageAppendix B: Table 4: Table of Cost Details of System ComponentsMd. Muhaiminul IslamNo ratings yet
- 4th Year Project ReportDocument2 pages4th Year Project ReportMd. Muhaiminul IslamNo ratings yet
- Diarrhoea, Is The Condition of Having at Least Three Loose or Liquid Bowel Movements Each Day. ItDocument6 pagesDiarrhoea, Is The Condition of Having at Least Three Loose or Liquid Bowel Movements Each Day. ItMd. Muhaiminul IslamNo ratings yet
- Abstract and Summary of Chapter 5Document1 pageAbstract and Summary of Chapter 5Md. Muhaiminul IslamNo ratings yet
- ANEMIADocument12 pagesANEMIAMd. Muhaiminul IslamNo ratings yet
- Definition:: Primary Gout: Secondary GoutDocument5 pagesDefinition:: Primary Gout: Secondary GoutMd. Muhaiminul IslamNo ratings yet
- Vitamin A DeficiencyDocument10 pagesVitamin A DeficiencyMd. Muhaiminul IslamNo ratings yet
- Iodine DeficiencyDocument10 pagesIodine DeficiencyMd. Muhaiminul IslamNo ratings yet
- Vitamin D CalciumDocument6 pagesVitamin D CalciumSajibNo ratings yet
- Protein Energy MalnutritionDocument19 pagesProtein Energy MalnutritionMd. Muhaiminul IslamNo ratings yet
- FTNS-3101 Nutritional Biochemistry-1: Rahanur AlamDocument22 pagesFTNS-3101 Nutritional Biochemistry-1: Rahanur AlamMd. Muhaiminul IslamNo ratings yet
- Iodine DeficiencyDocument10 pagesIodine DeficiencyMd. Muhaiminul IslamNo ratings yet
- ANEMIADocument12 pagesANEMIAMd. Muhaiminul IslamNo ratings yet
- Iodine DeficiencyDocument10 pagesIodine DeficiencyMd. Muhaiminul IslamNo ratings yet
- Diarrhoea, Is The Condition of Having at Least Three Loose or Liquid Bowel Movements Each Day. ItDocument6 pagesDiarrhoea, Is The Condition of Having at Least Three Loose or Liquid Bowel Movements Each Day. ItMd. Muhaiminul IslamNo ratings yet
- Definition:: Primary Gout: Secondary GoutDocument5 pagesDefinition:: Primary Gout: Secondary GoutMd. Muhaiminul IslamNo ratings yet
- FTNS-3101 Nutritional Biochemistry-1: Rahanur AlamDocument36 pagesFTNS-3101 Nutritional Biochemistry-1: Rahanur AlamMd. Muhaiminul IslamNo ratings yet
- Amino Acids OverviewDocument30 pagesAmino Acids OverviewbrownhazelNo ratings yet
- FTNS-3101 Nutritional Biochemistry-1: Rahanur AlamDocument80 pagesFTNS-3101 Nutritional Biochemistry-1: Rahanur AlamMd. Muhaiminul IslamNo ratings yet
- Lecture 03 PDFDocument4 pagesLecture 03 PDFMuhammad TariqNo ratings yet
- FTNS-3101-Oxidative PhosphorylationDocument4 pagesFTNS-3101-Oxidative PhosphorylationMd. Muhaiminul IslamNo ratings yet
- Amino AcidsDocument35 pagesAmino AcidsMd. Muhaiminul IslamNo ratings yet
- Amino Acids IIDocument64 pagesAmino Acids IIMd. Muhaiminul IslamNo ratings yet
- Ems em FW Paneel Firetec enDocument2 pagesEms em FW Paneel Firetec enzlatkokrsicNo ratings yet
- LapasiDocument3 pagesLapasiWenny MellanoNo ratings yet
- Allison Burke Adime 4Document8 pagesAllison Burke Adime 4api-317577095No ratings yet
- App Guide EntelliGuard - G 09 - 2020 AplicacionDocument100 pagesApp Guide EntelliGuard - G 09 - 2020 AplicacionjeorginagNo ratings yet
- 20BCEC1109, 20BCE1170, 20BCE1233 - IOT Final ReportDocument40 pages20BCEC1109, 20BCE1170, 20BCE1233 - IOT Final Reportharsh chauhanNo ratings yet
- Iso 2281 1990Document8 pagesIso 2281 1990jesus torresNo ratings yet
- Epilepsy 2 AAN 2018Document12 pagesEpilepsy 2 AAN 2018Eashaa KumarNo ratings yet
- Unsaturated HydrocarbonsDocument84 pagesUnsaturated HydrocarbonsHey itsJamNo ratings yet
- Keandalan Bangunan Rumah SusunDocument9 pagesKeandalan Bangunan Rumah SusunDewi ARimbiNo ratings yet
- BrainPOP Nutrition Quiz242342Document1 pageBrainPOP Nutrition Quiz242342MathableNo ratings yet
- DoveDocument11 pagesDovekattyperrysherryNo ratings yet
- June 2019Document64 pagesJune 2019Eric SantiagoNo ratings yet
- PNFDocument51 pagesPNFMuhamad Hakimi67% (3)
- Ethical Consideration in Leadership and ManagementDocument6 pagesEthical Consideration in Leadership and ManagementGlizzle Macaraeg67% (3)
- Open Courses Myanmar Strategic English Week 4: U Yan Naing Se NyuntDocument24 pagesOpen Courses Myanmar Strategic English Week 4: U Yan Naing Se NyuntYan Naing Soe NyuntNo ratings yet
- Osma Osmadrain BG Pim Od107 Feb 2017pdfDocument58 pagesOsma Osmadrain BG Pim Od107 Feb 2017pdfDeepakkumarNo ratings yet
- Canfield FairDocument3 pagesCanfield Fairapi-546463844No ratings yet
- Financial Markets & Institutions: Lecture Notes #3Document90 pagesFinancial Markets & Institutions: Lecture Notes #3Joan MaduNo ratings yet
- Chapter 3.2 Futures HedgingDocument19 pagesChapter 3.2 Futures HedginglelouchNo ratings yet
- The Importance of Early Childhood InterventionDocument11 pagesThe Importance of Early Childhood Interventionsilverlining0814100% (3)
- "Hiv and Aids Awareness of Junior and Senior High School Students" Part I: Profile of RespondentsDocument1 page"Hiv and Aids Awareness of Junior and Senior High School Students" Part I: Profile of RespondentsDahl xxNo ratings yet
- Basic Concepts and Applied Aspect of Sharir RachnaDocument122 pagesBasic Concepts and Applied Aspect of Sharir RachnaRam Krish100% (1)
- C50SCR PLC Complete ManualDocument286 pagesC50SCR PLC Complete ManualthuanNo ratings yet
- Evaluation of Whole-Body Vibration (WBV) On Ready Mixed Concrete Truck DriversDocument8 pagesEvaluation of Whole-Body Vibration (WBV) On Ready Mixed Concrete Truck DriversmariaNo ratings yet
- Work Environment Act (1977:1160) : Chapter 1 Purpose and Scope of The ActDocument26 pagesWork Environment Act (1977:1160) : Chapter 1 Purpose and Scope of The ActNatalie BlackmoreNo ratings yet
- MoringaDocument24 pagesMoringaobengappiahk100% (2)
- Creative An Inclusive ClassroomDocument3 pagesCreative An Inclusive Classroommuneeba zafarNo ratings yet
- Coiled Tubing Safety Impact Firing Head (HP) - by DynaDocument16 pagesCoiled Tubing Safety Impact Firing Head (HP) - by DynamahsaNo ratings yet
- Seachem Laboratories, Inc. Safety Data SheetDocument10 pagesSeachem Laboratories, Inc. Safety Data SheetJorge Restrepo HernandezNo ratings yet