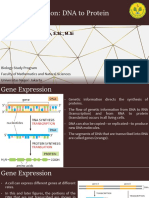
You might also like
- Biochemistry glossary termsDocument21 pagesBiochemistry glossary termsdivizioniNo ratings yet
- Study Dna DnaDocument45 pagesStudy Dna DnaKrystel BeltranNo ratings yet
- How Genes Work GENE - Biological Unit of HeredityDocument5 pagesHow Genes Work GENE - Biological Unit of HeredityAdonis Besa100% (1)
- Cell Biology Lecture NotesDocument108 pagesCell Biology Lecture NotesMert PolatNo ratings yet
- Dna Genes Chromosomes 2011Document65 pagesDna Genes Chromosomes 2011Suliman GarallehNo ratings yet
- Central Dogma of Molecular BiologyDocument30 pagesCentral Dogma of Molecular BiologyAlthea Mandal100% (1)
- 3rd Quarter W4 LECTURE On Protein Synthesis in DNA and MutationDocument53 pages3rd Quarter W4 LECTURE On Protein Synthesis in DNA and MutationMorante GeraldineNo ratings yet
- Inborn Errors of Metabolism LectureDocument32 pagesInborn Errors of Metabolism LectureEMMANUEL ABEL IMAH100% (1)
- In-Depth Steps Towards Nucleic Acid and Protein SynthesisDocument21 pagesIn-Depth Steps Towards Nucleic Acid and Protein SynthesisGbenga AjaniNo ratings yet
- Bio Exam ZsmuDocument84 pagesBio Exam ZsmuManthan Patil100% (1)
- Sintesis Protein TA 2023-2024Document111 pagesSintesis Protein TA 2023-2024fdla rhmahNo ratings yet
- Structure and Function of DNADocument29 pagesStructure and Function of DNAMatthew Justin Villanueva GozoNo ratings yet
- How Cells Work: The Central DogmaDocument18 pagesHow Cells Work: The Central DogmaJocelyn Diaz AbangNo ratings yet
- Aiatsoymeo2016t06 SolutionDocument29 pagesAiatsoymeo2016t06 Solutionsanthosh7kumar-24No ratings yet
- 6 BIO462 Chapter 6Document21 pages6 BIO462 Chapter 6Syafiqah SuhaimiNo ratings yet
- Usp 1116 em For Aseptic ProcessingDocument11 pagesUsp 1116 em For Aseptic ProcessingKraken Ur100% (1)
- Nucleic AcidsDocument37 pagesNucleic AcidsValine Cysteine MethionineNo ratings yet
- DNA's Structure and Function - CH 10Document10 pagesDNA's Structure and Function - CH 10jokersgang61No ratings yet
- Lecture 3 & 4: DNA, Genes, Chromatin Learning ObjectivesDocument65 pagesLecture 3 & 4: DNA, Genes, Chromatin Learning ObjectivesAli Al-QudsiNo ratings yet
- Structures and Functions of Nucleic AcidsDocument109 pagesStructures and Functions of Nucleic AcidsAisha A RehmanNo ratings yet
- CH1131 - Biomolecular Engineering - Week 2 - Transcription Translation - Aug 18 21 2015Document49 pagesCH1131 - Biomolecular Engineering - Week 2 - Transcription Translation - Aug 18 21 2015joshuaNo ratings yet
- DNA replication and protein synthesisDocument4 pagesDNA replication and protein synthesisKingNo ratings yet
- PROTEINS.123Document43 pagesPROTEINS.123agnasvynlutaoNo ratings yet
- DNA, RNA, and The Flow of InformationDocument31 pagesDNA, RNA, and The Flow of InformationSasa AbassNo ratings yet
- Paper - IV Molecular BiologyDocument236 pagesPaper - IV Molecular BiologyNaruto UzumakiNo ratings yet
- Dna, Rna, & Proteins DNA Replication and Protein Synthesis: Central Dogma of Molecular BiologyDocument79 pagesDna, Rna, & Proteins DNA Replication and Protein Synthesis: Central Dogma of Molecular BiologynokeNo ratings yet
- Module 8 - Nucleic Acids and Central DogmaDocument52 pagesModule 8 - Nucleic Acids and Central DogmaPauline Grace CadusaleNo ratings yet
- Information Flow From DNA To Protein - Lecture SlidesDocument66 pagesInformation Flow From DNA To Protein - Lecture Slidesjudssssk46No ratings yet
- Ekspresi Gen - Bruce Albert (SEND)Document14 pagesEkspresi Gen - Bruce Albert (SEND)Rahmah Khairunnisa QonitaNo ratings yet
- Nucleic Acids SummaryDocument4 pagesNucleic Acids SummaryIvan MiglioriniNo ratings yet
- Chapter 4A: DNA Replication & Protein SynthesisDocument34 pagesChapter 4A: DNA Replication & Protein SynthesisPikuNo ratings yet
- 6.2. DNA Replication PDFDocument75 pages6.2. DNA Replication PDFShania TehNo ratings yet
- DNA and RNA: The Molecules of HeredityDocument42 pagesDNA and RNA: The Molecules of HeredityLyssaMarieKathryneEgeNo ratings yet
- Structure and Function of DNADocument27 pagesStructure and Function of DNAHaniya RijaNo ratings yet
- Basics of Molecular BiologyDocument10 pagesBasics of Molecular BiologyOday MadiNo ratings yet
- DNA Replication and the Central DogmaDocument94 pagesDNA Replication and the Central Dogmameklit birhanuNo ratings yet
- Unit 7 Nucleic AcidsDocument62 pagesUnit 7 Nucleic AcidsGlory MuedaNo ratings yet
- Genetic Makeup and Replication of Bacterial Cell MAA-1Document37 pagesGenetic Makeup and Replication of Bacterial Cell MAA-1she.dil.oneNo ratings yet
- Week 5 - GeneticsDocument23 pagesWeek 5 - GeneticsJuan Pablo Valencia LondoñoNo ratings yet
- Structure of DNA Science Presentation in Light Blue Green Lined Style 20240314 1Document26 pagesStructure of DNA Science Presentation in Light Blue Green Lined Style 20240314 1leighariazbongsNo ratings yet
- Structure and Functions of DNADocument37 pagesStructure and Functions of DNARaj kumar paswanNo ratings yet
- Nucleotides and Nucleic AcidsDocument68 pagesNucleotides and Nucleic AcidsVaibhav KumarNo ratings yet
- Eukaryotic GenomesDocument50 pagesEukaryotic GenomesturkiNo ratings yet
- Lesson-3 Protein-Synthesis WorksheetDocument10 pagesLesson-3 Protein-Synthesis WorksheetEnriquez, Hannah Roniella R.No ratings yet
- Replikasi Dna Repair Dna Rekombinasi Dna: Dr. Upik A. Miskad, PHD Unhas Research CenterDocument48 pagesReplikasi Dna Repair Dna Rekombinasi Dna: Dr. Upik A. Miskad, PHD Unhas Research CenterFitriany LihawaNo ratings yet
- 2 Dna Structure and RnaDocument110 pages2 Dna Structure and Rnakhan bhaiNo ratings yet
- Biology: Third QuarterDocument126 pagesBiology: Third QuarterHill A. D.No ratings yet
- The Central Dogma of A Cell: DR S.Balakrishnan, PHD.Document58 pagesThe Central Dogma of A Cell: DR S.Balakrishnan, PHD.joyousNo ratings yet
- General Biology 2Document4 pagesGeneral Biology 2Christianne Joseph MonzonNo ratings yet
- Designer Genes: Practice - Molecular-Genetic GeneticsDocument26 pagesDesigner Genes: Practice - Molecular-Genetic GeneticsGiovanni TorresNo ratings yet
- Molecular Biology: BBM FK UntarDocument47 pagesMolecular Biology: BBM FK UntarfaustineNo ratings yet
- Nucleotides and Nucleic AcidsDocument65 pagesNucleotides and Nucleic AcidsnutriNo ratings yet
- DNA Structure and Replication ExplainedDocument37 pagesDNA Structure and Replication ExplainedJohn KennedyNo ratings yet
- Biological BackgroundDocument15 pagesBiological Backgroundrustyryan77No ratings yet
- Replication Transcription and TranslationDocument16 pagesReplication Transcription and TranslationBalangat Regine L.No ratings yet
- Genetics: Unit III - Living Things and Its Environment - Heredity: Inheritance & VariationDocument4 pagesGenetics: Unit III - Living Things and Its Environment - Heredity: Inheritance & VariationKirito TobioNo ratings yet
- Chapter 12 Biol1010 Notes-1-1Document4 pagesChapter 12 Biol1010 Notes-1-1yazst.julienNo ratings yet
- DNA Structure and ReplicationDocument30 pagesDNA Structure and ReplicationasaadsarfrazNo ratings yet
- DNA Replication in Molecular BiologyDocument9 pagesDNA Replication in Molecular BiologyАбу ДжудNo ratings yet
- Central Dogma: Dna ReplicationDocument18 pagesCentral Dogma: Dna ReplicationAkhil KumarNo ratings yet
- 6 - Microbial GeneticsDocument35 pages6 - Microbial GeneticsAbdallah MoayadNo ratings yet
- BCH 401 Note 1 - KWASUDocument35 pagesBCH 401 Note 1 - KWASUidriscognitoleadsNo ratings yet
- Lavaughn's DNA, RNA PresentationDocument12 pagesLavaughn's DNA, RNA Presentationpnjq6sjrx7No ratings yet
- Module 4 KKKDocument32 pagesModule 4 KKKbhrayancachero100% (1)
- BLG413 Chapter 1 Part 2Document19 pagesBLG413 Chapter 1 Part 2rmf92r4wbrNo ratings yet
- 5 - Amino AcidsDocument12 pages5 - Amino AcidsEMMANUEL ABEL IMAHNo ratings yet
- TCA CycleDocument22 pagesTCA CycleEMMANUEL ABEL IMAHNo ratings yet
- Blood SmearDocument15 pagesBlood SmearEMMANUEL ABEL IMAHNo ratings yet
- BMD 300 Pasco. Prof Cosmos LibraryDocument53 pagesBMD 300 Pasco. Prof Cosmos LibraryEMMANUEL ABEL IMAHNo ratings yet
- "They Will Fight Against You But Will Not Overcome You, For I Am With You and Will Rescue You," Declares The LORDDocument76 pages"They Will Fight Against You But Will Not Overcome You, For I Am With You and Will Rescue You," Declares The LORDemmanuel imahNo ratings yet
- MBB425 - Lecture 2-3 GENOME STRUCTUREDocument35 pagesMBB425 - Lecture 2-3 GENOME STRUCTUREEMMANUEL ABEL IMAHNo ratings yet
- Lecture9 Genetic Code & Protein SynthesisDocument64 pagesLecture9 Genetic Code & Protein SynthesisConstance WongNo ratings yet
- MSC BiotechnologyDocument82 pagesMSC BiotechnologyJanikaa Singaravel MuruganNo ratings yet
- SCIENCEDocument17 pagesSCIENCECJMDNo ratings yet
- Microbial Evolution and SystematicsDocument76 pagesMicrobial Evolution and SystematicsDom GudezNo ratings yet
- BiochemDocument383 pagesBiochemtylermedNo ratings yet
- HesterDocument44 pagesHesterDyah tantri WNo ratings yet
- Landers 2020Document10 pagesLanders 2020Abraham Flores RendónNo ratings yet
- Nucleic AcidsDocument56 pagesNucleic AcidsShane G.No ratings yet
- Taha 2017Document9 pagesTaha 2017aufa QooNo ratings yet
- Introducción - La CélulaDocument35 pagesIntroducción - La CélulaVelia Lopez SanchezNo ratings yet
- Paper I Unit Ii:-Gene Expression Rna Polymerase DefinitionDocument53 pagesPaper I Unit Ii:-Gene Expression Rna Polymerase DefinitionRithwik ZNo ratings yet
- RN AmmerDocument2 pagesRN Ammerbioefl33No ratings yet
- Translation: The Process of Protein SynthesisDocument12 pagesTranslation: The Process of Protein SynthesisHarshit YadavNo ratings yet
- Evolution Evidence ExplainedDocument13 pagesEvolution Evidence ExplainedJohn Bernard VeloriaNo ratings yet
- Module 3Document15 pagesModule 3RabbitFamNo ratings yet
- Molecular Microbial Diversity of An Agricultural Soil in WisconsinDocument9 pagesMolecular Microbial Diversity of An Agricultural Soil in Wisconsinshailu140287No ratings yet
- Culture BacteriaDocument6 pagesCulture BacteriaCelina PereiraNo ratings yet
- Bio 111 Final Exam Fall05Document21 pagesBio 111 Final Exam Fall05Zeshan AslamNo ratings yet
- Molecular BiologyDocument20 pagesMolecular BiologyDaphne CuaresmaNo ratings yet
- Zhao 2017Document10 pagesZhao 2017LUIS GIANFRANCO GONZALEZ ALVAREZNo ratings yet
- Formula Sheet Molecular Basis of Inheritance 1Document6 pagesFormula Sheet Molecular Basis of Inheritance 1Aarthi T. UNo ratings yet
- Prokaryotic and Eukaryotic CellsDocument24 pagesProkaryotic and Eukaryotic Cells2begeniusNo ratings yet
- Topic 10 Transcription Lecture NotesDocument33 pagesTopic 10 Transcription Lecture NoteskambulukatambilaiNo ratings yet
- Bulu BabiDocument17 pagesBulu BabiAswad AffandiNo ratings yet
- Monday Test 7 SolutionDocument11 pagesMonday Test 7 Solutionbiotecnika_test100% (1)