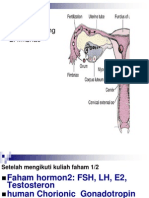
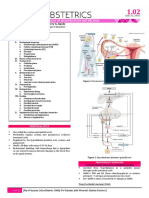
You might also like
- Normal Menstrual CycleDocument28 pagesNormal Menstrual CycleCristóbal ConchaNo ratings yet
- Menstrual Cycle AutosavedDocument33 pagesMenstrual Cycle Autosavedნინო ღვინაძე100% (1)
- Menstrual Irregularities: Causes and Tracking ChangesDocument6 pagesMenstrual Irregularities: Causes and Tracking ChangesLioraNo ratings yet
- The Menstrual Cycle Ob - Gyn SecretsDocument4 pagesThe Menstrual Cycle Ob - Gyn Secretsleonardotorres1No ratings yet
- TP Anglais FinalDocument7 pagesTP Anglais FinalGift NgongoNo ratings yet
- MCN Cover To Cover ?Document247 pagesMCN Cover To Cover ?Elsid Nathaniel S. MartinNo ratings yet
- Topic-Menstural Cycle: Name-Divya Sharma Roll No-8 & Bharti Kotwal Roll No-7Document15 pagesTopic-Menstural Cycle: Name-Divya Sharma Roll No-8 & Bharti Kotwal Roll No-7chanderNo ratings yet
- Menstrual CycleDocument24 pagesMenstrual Cyclemeez rameezNo ratings yet
- Physiology of Women Reproduction SystemDocument52 pagesPhysiology of Women Reproduction Systemram kumarNo ratings yet
- IntroDocument7 pagesIntroibrahim.21hm32No ratings yet
- Physiology of The Normal Menstrual Cycle - UpToDateDocument14 pagesPhysiology of The Normal Menstrual Cycle - UpToDatefranciscaNo ratings yet
- The Ovarian Cycle, The Menstrual Cycle, and MenopauseDocument1 pageThe Ovarian Cycle, The Menstrual Cycle, and MenopauseMohamad Hilman Al-KelantaniNo ratings yet
- 1642497170754_Family Health 1 - CopyDocument20 pages1642497170754_Family Health 1 - CopyGoogle MediaNo ratings yet
- ILA4Document7 pagesILA4Fadhlina OmarNo ratings yet
- The Menstrual Cycle TranskipDocument5 pagesThe Menstrual Cycle Transkiplen lehangNo ratings yet
- Problem No.1Document34 pagesProblem No.1ayesha bgNo ratings yet
- Hormonal Cycles: Presented By: Miss. M.K.Kaku Nursing TutorDocument23 pagesHormonal Cycles: Presented By: Miss. M.K.Kaku Nursing TutorKaku ManishaNo ratings yet
- HORMONAL CYCLE EXPLAINEDDocument20 pagesHORMONAL CYCLE EXPLAINEDGunaNo ratings yet
- Female Physiology 1Document2 pagesFemale Physiology 1JayricDepalobosNo ratings yet
- Chapter 9 - Disorders Related To Menstruation PDFDocument31 pagesChapter 9 - Disorders Related To Menstruation PDFLidia CampeanNo ratings yet
- Reproductive Cycles in MammalsDocument9 pagesReproductive Cycles in MammalsSeemal shahidNo ratings yet
- The Menstrual Cycle P. Mock: Infertility and Gynecologic Endocrinology Clinic Geneva University HospitalDocument12 pagesThe Menstrual Cycle P. Mock: Infertility and Gynecologic Endocrinology Clinic Geneva University HospitalMubashir's iPhoneNo ratings yet
- Menstrual Cycle HormonesDocument3 pagesMenstrual Cycle HormonesAine GrahamNo ratings yet
- Normal Menstrual Cycle The ProcessDocument6 pagesNormal Menstrual Cycle The ProcessBrett StevensonNo ratings yet
- Hormonal Cycles: Ovarian CycleDocument5 pagesHormonal Cycles: Ovarian CycleashuNo ratings yet
- Menstrual Cycle Biology ProjectDocument4 pagesMenstrual Cycle Biology ProjectaddyNo ratings yet
- Menstrual CycleDocument66 pagesMenstrual CycleMelizza Fajardo Bañano100% (1)
- Q3 Lesson 3 The Female Reproductive Feedback MechanismDocument27 pagesQ3 Lesson 3 The Female Reproductive Feedback Mechanismjademarco901No ratings yet
- Menstrual CycleDocument41 pagesMenstrual CyclesharimileeratnamNo ratings yet
- The Menstrual Cycle by CK-12Document7 pagesThe Menstrual Cycle by CK-12woman in stemNo ratings yet
- Physiology of Female Reproductive System The Ovarian CycleDocument9 pagesPhysiology of Female Reproductive System The Ovarian CycleAmiraah MasriNo ratings yet
- MCN Menstrual Cycle LatestDocument8 pagesMCN Menstrual Cycle LatestsophienNo ratings yet
- Menstrual Cycle Hormones ExplainedDocument29 pagesMenstrual Cycle Hormones ExplainedLiezel CauilanNo ratings yet
- Meenal BiologyDocument18 pagesMeenal BiologymansiNo ratings yet
- Menstrual Cycle FinalDocument28 pagesMenstrual Cycle FinalMary Claire LumbaoNo ratings yet
- Self-Learning Home Task (SLHT)Document9 pagesSelf-Learning Home Task (SLHT)GraceEstoleCaloNo ratings yet
- 1 Normal Ob (Menstrual Cycle) : Glora P. de Leon, RN, RM, ManDocument75 pages1 Normal Ob (Menstrual Cycle) : Glora P. de Leon, RN, RM, ManJoan VillafrancaNo ratings yet
- Female Reproductive CycleDocument4 pagesFemale Reproductive CycleTRASH MAILNo ratings yet
- Chapter 2 - Physiology of MenstruationDocument18 pagesChapter 2 - Physiology of Menstruationاسامة محمد السيد رمضان100% (1)
- Reproductive SystemDocument30 pagesReproductive SystemmfaizchejamriNo ratings yet
- Female HormoneDocument43 pagesFemale Hormoneامجد حسين جواد كاظمNo ratings yet
- Unit 3 Module 1 The Menstrual CycleDocument20 pagesUnit 3 Module 1 The Menstrual CycleMARIANNE SORIANO100% (2)
- Unit VIII Physiology of Pregnancy (Changes and Adaptation)Document10 pagesUnit VIII Physiology of Pregnancy (Changes and Adaptation)Binita ShakyaNo ratings yet
- MK Disorders of Menstruation (OBGY)Document76 pagesMK Disorders of Menstruation (OBGY)Moses Jr KazevuNo ratings yet
- Understanding the Menstrual CycleDocument10 pagesUnderstanding the Menstrual CyclebfgbffdNo ratings yet
- Notes Infertility Delayed PubertyDocument9 pagesNotes Infertility Delayed PubertyTONY GO AWAYNo ratings yet
- Menstrual CycleDocument14 pagesMenstrual CycleVáncsa Szilárd100% (2)
- CycleDocument2 pagesCycleChinchay Guinto100% (1)
- Menstrual CycleDocument4 pagesMenstrual CycleIce-ice Valencia-SulaikNo ratings yet
- Female Repoductive CyclesDocument11 pagesFemale Repoductive CyclesbokkNo ratings yet
- Imported FileDocument18 pagesImported Fileg8pn6pt8nhNo ratings yet
- Puberty Below: Role of HormonesDocument3 pagesPuberty Below: Role of HormonesCyril CauilanNo ratings yet
- Investigatory Project of BiologyDocument18 pagesInvestigatory Project of BiologyRanjana Singh87% (31)
- 4 Endocrine 6Document16 pages4 Endocrine 6ShenNo ratings yet
- Menstrual Cycle: An IntroductionDocument6 pagesMenstrual Cycle: An IntroductionAudrie Allyson GabalesNo ratings yet
- Physiology of the Menstrual CycleDocument6 pagesPhysiology of the Menstrual CyclegmindalanoNo ratings yet
- Kuliah PK2 TESKehamilanJULI2010Document38 pagesKuliah PK2 TESKehamilanJULI2010Fetty NataliaNo ratings yet
- Study Session 4 Hormonal Regulation of The Female Reproductive SystemDocument11 pagesStudy Session 4 Hormonal Regulation of The Female Reproductive SystemUMINAHNo ratings yet
- Hormone Reset Diet: Restore Your Metabolism, Sex Drive and Get Your Life Back, All While Losing 15lbsFrom EverandHormone Reset Diet: Restore Your Metabolism, Sex Drive and Get Your Life Back, All While Losing 15lbsNo ratings yet
- ch-6 of Science Class 6Document4 pagesch-6 of Science Class 6sanjeev kumarNo ratings yet
- Forests: Our Lifeline: Presented By: Subhash Kumar Class, Section: VII D Roll No:42Document5 pagesForests: Our Lifeline: Presented By: Subhash Kumar Class, Section: VII D Roll No:42sanjeev kumarNo ratings yet
- Presentation of An Earthquake: From Name Subhash Kumar Class VII D Roll No 42Document5 pagesPresentation of An Earthquake: From Name Subhash Kumar Class VII D Roll No 42sanjeev kumarNo ratings yet
- Madhubani Paintings: From Subhash Kumar Class Viith D Roll NoDocument4 pagesMadhubani Paintings: From Subhash Kumar Class Viith D Roll Nosanjeev kumarNo ratings yet
- Infertility in MaresDocument7 pagesInfertility in MaresYaserAbbasiNo ratings yet
- LIFE SCIENCES Grade 12 Trial 2021 QP and MemoDocument25 pagesLIFE SCIENCES Grade 12 Trial 2021 QP and MemoDonoven Malotane100% (1)
- Maternal and Child Health Is The Program of The Department of Health Created To Lessen The Death of Infants and Mother in The PhilippinesDocument18 pagesMaternal and Child Health Is The Program of The Department of Health Created To Lessen The Death of Infants and Mother in The PhilippinesNicole Marin-ChingNo ratings yet
- Lecture 2-Pregnancy and Parturition in Animals 2019-1Document10 pagesLecture 2-Pregnancy and Parturition in Animals 2019-1Winnie PillyNo ratings yet
- Estrogen Dominance Part 2 TherapeuticsDocument34 pagesEstrogen Dominance Part 2 TherapeuticsAnthony De PontesNo ratings yet
- 1638366605-EASE 2 Pointers - Grade 8Document4 pages1638366605-EASE 2 Pointers - Grade 8Zian hafisNo ratings yet
- Disorders of The Ovaries 2Document92 pagesDisorders of The Ovaries 2rerenNo ratings yet
- Sleep and Disorders of Sleep in Women, An Issue of Sleep Medicine ClinicsDocument138 pagesSleep and Disorders of Sleep in Women, An Issue of Sleep Medicine ClinicsPatrickNo ratings yet
- Government College of Nursing:, Jodhpur (RAJ.)Document12 pagesGovernment College of Nursing:, Jodhpur (RAJ.)priyanka100% (1)
- Chapter 18 Biology 2nd Year - Prof. Ijaz Ahmed Khan Abbasi (Lecturer Biology PGC) Notes - MDCAT by FUTURE DOCTORS - Touseef Ahmad Khan - 03499815886Document25 pagesChapter 18 Biology 2nd Year - Prof. Ijaz Ahmed Khan Abbasi (Lecturer Biology PGC) Notes - MDCAT by FUTURE DOCTORS - Touseef Ahmad Khan - 03499815886Suffa AcademyNo ratings yet
- The Test May Be Run On Patients Between 22 and 34 Weeks GestationDocument6 pagesThe Test May Be Run On Patients Between 22 and 34 Weeks Gestationdaniel_alexander_susenoNo ratings yet
- Effects of Follicular and Luteal Phase Based Menstrual Cycle Resistance Training On Muscle Strength and MassDocument7 pagesEffects of Follicular and Luteal Phase Based Menstrual Cycle Resistance Training On Muscle Strength and MassEmily ClarkNo ratings yet
- Family Medicine & Medical Science Research: Cabergoline (Dostinex) Effect On Weight Ovarian FunctionDocument3 pagesFamily Medicine & Medical Science Research: Cabergoline (Dostinex) Effect On Weight Ovarian FunctionAbo Marwan AlhomsiNo ratings yet
- Long Quiz MCN LecDocument3 pagesLong Quiz MCN LecShing Mae MarieNo ratings yet
- Feedback Mechanisms and Menstrual CycleDocument90 pagesFeedback Mechanisms and Menstrual CycleThird WillowNo ratings yet
- Menstrual Cycle Alters Face PreferenceDocument2 pagesMenstrual Cycle Alters Face PreferenceInterestingScienceNo ratings yet
- Obstetrics and Gynecology MCQsDocument34 pagesObstetrics and Gynecology MCQsARUSHI ARVINDNo ratings yet
- BIO ProjectDocument17 pagesBIO ProjectHârsh V ÎshwãkårmāNo ratings yet
- DLL Sci Q2 W2Document12 pagesDLL Sci Q2 W2Raymund DelfinNo ratings yet
- OB 1 - 1.02 Physiology of Menstruation and Decidua PDFDocument11 pagesOB 1 - 1.02 Physiology of Menstruation and Decidua PDFMDreamerNo ratings yet
- Dharumavantha School Examinations: First Term 2009 Grade 10Document11 pagesDharumavantha School Examinations: First Term 2009 Grade 10afoo1234No ratings yet
- VGO-411 Veterinary Gynaecology Question BankDocument7 pagesVGO-411 Veterinary Gynaecology Question BankSuryakantaRoulTuntunNo ratings yet
- FertilityDocument207 pagesFertilitypmotcNo ratings yet
- Physiology of MenstruationDocument52 pagesPhysiology of Menstruationmahirad001100% (2)
- Embryo Transfer TechnologyDocument55 pagesEmbryo Transfer TechnologyShubham AgrawalNo ratings yet
- Set 3 - Diagnostic TestDocument14 pagesSet 3 - Diagnostic Testdanielkenneth deleñaNo ratings yet
- Fetal Development Month by Month Stages of PregnancyDocument11 pagesFetal Development Month by Month Stages of PregnancyBea YmsnNo ratings yet
- Compare and Contrast Plant and Animal Reproduction and DevelopmentDocument145 pagesCompare and Contrast Plant and Animal Reproduction and DevelopmentAuxidyNo ratings yet
- Understanding AnovulationDocument207 pagesUnderstanding AnovulationRosu George100% (2)
- Recent Advances in Animal Nutrition 2009Document344 pagesRecent Advances in Animal Nutrition 2009Juan NfrNo ratings yet