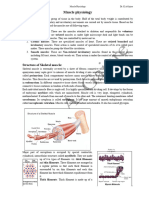
You might also like
- Skeletal MuscleDocument98 pagesSkeletal MuscleMoon Moon100% (1)
- Muscle Physiology in Orthodontics / Orthodontic Courses by Indian Dental AcademyDocument50 pagesMuscle Physiology in Orthodontics / Orthodontic Courses by Indian Dental Academyindian dental academyNo ratings yet
- The Basics of Cell Life with Max Axiom, Super Scientist: 4D An Augmented Reading Science ExperienceFrom EverandThe Basics of Cell Life with Max Axiom, Super Scientist: 4D An Augmented Reading Science ExperienceNo ratings yet
- HIV - PathogenesisDocument14 pagesHIV - Pathogenesisjunaidiabdhalim100% (1)
- Physiology of Muscle Contraction and Control of MovementDocument69 pagesPhysiology of Muscle Contraction and Control of MovementMakmunNawil100% (1)
- Intro To Biohacking: or "How I Learned To Stop Worrying and Love The Zombie Apocalypse"Document31 pagesIntro To Biohacking: or "How I Learned To Stop Worrying and Love The Zombie Apocalypse"rasromeoNo ratings yet
- Microscopic Anatomy of A Skeletal Muscle FiberDocument33 pagesMicroscopic Anatomy of A Skeletal Muscle FiberKumar MSNo ratings yet
- Wa0038Document10 pagesWa0038Shehzad QureshiNo ratings yet
- General Mechanism of Muscle ContractionDocument22 pagesGeneral Mechanism of Muscle ContractionAnuya Kulkarni-DeoNo ratings yet
- Physiology of Skeletal & Smooth MusclesDocument10 pagesPhysiology of Skeletal & Smooth MusclesAli AlhamadNo ratings yet
- Muscles MidtermDocument32 pagesMuscles MidtermMata, Ersel MaeNo ratings yet
- Muscular SystemDocument10 pagesMuscular SystemMarie Veatrice JacomilleNo ratings yet
- Revision Notes on Locomotion and Movement for NEET 2024 - Free PDF DownloadDocument11 pagesRevision Notes on Locomotion and Movement for NEET 2024 - Free PDF DownloadHaider KaziNo ratings yet
- (L2) - Locomotion and Movement - Sept 25, 2019 - WednesdayDocument45 pages(L2) - Locomotion and Movement - Sept 25, 2019 - WednesdayRofiul HussainNo ratings yet
- ST - of Myofilaments..Document10 pagesST - of Myofilaments..Mohammed HamadaNo ratings yet
- Chapter 6Document2 pagesChapter 6Nona PeriarceNo ratings yet
- The Muscular SystemDocument9 pagesThe Muscular SystemKathleenJoyGalAlmasinNo ratings yet
- B Muscle Tissue HistologyDocument55 pagesB Muscle Tissue HistologyAffie SaikolNo ratings yet
- Muscle Lecture On (Excitable Tissue)Document126 pagesMuscle Lecture On (Excitable Tissue)Muhammed ZainabNo ratings yet
- Human Anatomy and Physiology - Structure and Mechanism of Skeletal Muscle ContractionDocument14 pagesHuman Anatomy and Physiology - Structure and Mechanism of Skeletal Muscle ContractionRAI SUNNo ratings yet
- Actin Myosin CytoskeletonDocument28 pagesActin Myosin Cytoskeletonjesus rajeshdivyaNo ratings yet
- SM Skeletal Muscle SlidesDocument68 pagesSM Skeletal Muscle Slideskarishma.7022No ratings yet
- Locomotion and MovementDocument9 pagesLocomotion and Movementpoojari.dedeepyaNo ratings yet
- Physiology Slide #1Document18 pagesPhysiology Slide #1anon_974423120No ratings yet
- Trabalho BioquimicaDocument12 pagesTrabalho BioquimicaRicardo PessoaNo ratings yet
- Chapter VI - Contraction of The Skeletal Muscle: Filamentous Bands Light Bands Dark BandsDocument13 pagesChapter VI - Contraction of The Skeletal Muscle: Filamentous Bands Light Bands Dark BandsLorenz L. Llamas IIINo ratings yet
- Chapter 9 Actin FloresDocument39 pagesChapter 9 Actin FloresPATRICK MARK FLORESNo ratings yet
- Muscle PhysiologyDocument12 pagesMuscle PhysiologyAyres EvangNo ratings yet
- 589bc774 Df1d 47c6 Bc97 b5fd73030ca6Muscle PhysiologyDocument3 pages589bc774 Df1d 47c6 Bc97 b5fd73030ca6Muscle PhysiologyMalti SharmaNo ratings yet
- 6.muscular TissueDocument40 pages6.muscular Tissuemero wowNo ratings yet
- Skeletal MusclesDocument7 pagesSkeletal MusclesAhmed OudahNo ratings yet
- Muscle ContractionDocument21 pagesMuscle ContractionMarilyn GumeyiNo ratings yet
- 3D. Muscle TissueDocument47 pages3D. Muscle Tissuejaf123xyNo ratings yet
- MuscleDocument11 pagesMuscleJeff ParkNo ratings yet
- L 9 MyosinDocument22 pagesL 9 Myosindiksha singhNo ratings yet
- Physiology of Skeletal Muscle and Muscle FibreDocument7 pagesPhysiology of Skeletal Muscle and Muscle FibreNoor-E-Khadiza ShamaNo ratings yet
- Muscular System FunctionsDocument17 pagesMuscular System FunctionsNicoleVideñaNo ratings yet
- Reviewer - Muscular SystemDocument11 pagesReviewer - Muscular SystemIvy Jan OcateNo ratings yet
- Muscle PhysiologyDocument24 pagesMuscle PhysiologyArmando AlehandroNo ratings yet
- Skeletal Muscle PDFDocument45 pagesSkeletal Muscle PDFyusufalper7234No ratings yet
- Faal Otot Dan GerakDocument46 pagesFaal Otot Dan GerakElsya Nur FadhillaNo ratings yet
- Chapter 7 - Muscular SystemDocument29 pagesChapter 7 - Muscular SystemlNo ratings yet
- Locomotion & Movement PowernotesDocument3 pagesLocomotion & Movement PowernotesSushmit Srivastava100% (2)
- CytoskeletonDocument27 pagesCytoskeletonserenity limNo ratings yet
- Muscle DentalDocument74 pagesMuscle Dentalapi-3710331No ratings yet
- Chapter 5Document25 pagesChapter 5أمال داودNo ratings yet
- Movement Biopsych NotesDocument18 pagesMovement Biopsych NotesRealyn ZambasNo ratings yet
- Muscle PhysiologyDocument88 pagesMuscle PhysiologyAftab MughalNo ratings yet
- Bio 215 Topic 5 MusclesDocument88 pagesBio 215 Topic 5 MusclesYara BeainiNo ratings yet
- Skeletal Muscle Classification and StructureDocument43 pagesSkeletal Muscle Classification and StructureHusnain WattoNo ratings yet
- Understanding Muscle PhysiologyDocument88 pagesUnderstanding Muscle Physiologyandre andreNo ratings yet
- Skeletal Muscle Physiology ExplainedDocument26 pagesSkeletal Muscle Physiology ExplainedUSAMA AHMEDNo ratings yet
- Muscular TDocument18 pagesMuscular TSumayya KabeerNo ratings yet
- Muscle Tissue: - Skeletal Muscle - Cardiac Muscle - Smooth MuscleDocument36 pagesMuscle Tissue: - Skeletal Muscle - Cardiac Muscle - Smooth MuscleDentist Dina Samy100% (1)
- Unit 3 Muscle Physiology - BPT PDFDocument55 pagesUnit 3 Muscle Physiology - BPT PDFRadhikaNo ratings yet
- Locomotion & Movement (Muscles)Document41 pagesLocomotion & Movement (Muscles)DEEPANKAR SAHANo ratings yet
- MuscleDocument5 pagesMuscleAbdulrahman AqraNo ratings yet
- Muscular SystemDocument9 pagesMuscular SystemReynaldo MontañoNo ratings yet
- Muscles1Document25 pagesMuscles1sairash.khan23No ratings yet
- Structre of Skeletal MuscleDocument14 pagesStructre of Skeletal MuscleAdam PrabowoNo ratings yet
- Anaphy-Muscular HandoutDocument7 pagesAnaphy-Muscular HandoutAEGEL DARO TORRALBANo ratings yet
- Final Paper 1Document140 pagesFinal Paper 1Ramil LucasNo ratings yet
- Bone Key 2012183Document8 pagesBone Key 2012183Jesus MendezNo ratings yet
- SMK Petaling Test Specification Table (Jsu) Biology Form 4 Paper 2Document2 pagesSMK Petaling Test Specification Table (Jsu) Biology Form 4 Paper 2Raidah AdhamNo ratings yet
- Experiment 3 - Graphite PhagocytosisDocument26 pagesExperiment 3 - Graphite PhagocytosisKristella Draheim0% (1)
- The Role of Dimethylaminoethanol in Cosmetic Dermatology: Rachel GrossmanDocument26 pagesThe Role of Dimethylaminoethanol in Cosmetic Dermatology: Rachel GrossmanLibe Palomino RivasNo ratings yet
- Unique Properties of All Stem Cells PDFDocument1 pageUnique Properties of All Stem Cells PDFExicanLifeSciencesNo ratings yet
- Lesson 1 Cellular Processes PPTDocument19 pagesLesson 1 Cellular Processes PPTkj47ynpdh7No ratings yet
- The New Biology and Pharmacology of GlucagonDocument46 pagesThe New Biology and Pharmacology of GlucagonAntonio SalemeNo ratings yet
- This Study Resource Was: DNA Base Pairing WorksheetDocument5 pagesThis Study Resource Was: DNA Base Pairing Worksheetphil tolentinoNo ratings yet
- Gizmo - Rna - Protein SynthesisDocument7 pagesGizmo - Rna - Protein SynthesisMarques AlsoppNo ratings yet
- Tema 7quim Al17-18Document53 pagesTema 7quim Al17-18noelia100997No ratings yet
- Amali Alternatif Experiment 3 (Prosedur Alternatif)Document4 pagesAmali Alternatif Experiment 3 (Prosedur Alternatif)Jennie RosèNo ratings yet
- Avanti Brochure PEG LipidsDocument2 pagesAvanti Brochure PEG LipidspalkybdNo ratings yet
- AP+Bio 042+Biological+Molecules+Worksheet WLDocument2 pagesAP+Bio 042+Biological+Molecules+Worksheet WLSabrina LouiseNo ratings yet
- Notch-Activated Signaling Cascade Interacts With Mitochondrial Remodeling Proteins To Regulate Cell SurvivalDocument6 pagesNotch-Activated Signaling Cascade Interacts With Mitochondrial Remodeling Proteins To Regulate Cell SurvivalFarah HaqueNo ratings yet
- 3.1 Cell Structure Ans PDFDocument6 pages3.1 Cell Structure Ans PDFtess_15No ratings yet
- JEM DCs Macrophages Special IssueDocument76 pagesJEM DCs Macrophages Special IssueAbdurRahmanNo ratings yet
- Wilson - PhysioEx Exercise 1 Activity 5Document3 pagesWilson - PhysioEx Exercise 1 Activity 5Hulwah ShalihahNo ratings yet
- Coutu 2017Document12 pagesCoutu 2017ThunderslapNo ratings yet
- Lab Report 2 Final - Botany1Document9 pagesLab Report 2 Final - Botany1Jonathan SerranoNo ratings yet
- 13 Photosynthesis: Key TermsDocument7 pages13 Photosynthesis: Key Termssalahuddin_md5935No ratings yet
- Embryo AssignmentDocument2 pagesEmbryo AssignmentA.j. MasagcaNo ratings yet
- Citric Acid Cycle Enzyme LabDocument9 pagesCitric Acid Cycle Enzyme LabDan Pingel100% (1)
- Concise Histology Gartner, Leslie PDFDocument478 pagesConcise Histology Gartner, Leslie PDFLexsshNo ratings yet
- Chapter 9 Cardiac MuscleDocument5 pagesChapter 9 Cardiac MuscleYheng GaosaiiNo ratings yet
- Gram Staining TechniqueDocument13 pagesGram Staining TechniqueKing Carlos FriasNo ratings yet
- CHM4116 Nitrogen Metab ReviewerDocument22 pagesCHM4116 Nitrogen Metab ReviewerBEA FRANCINE DELOS SANTOSNo ratings yet
- Malignant Peripheral Nerve Sheath Tumors: Differentiation Patterns and Immunohistochemical Features - A Mini-Review and Our New FindingsDocument7 pagesMalignant Peripheral Nerve Sheath Tumors: Differentiation Patterns and Immunohistochemical Features - A Mini-Review and Our New FindingsAfkar30No ratings yet