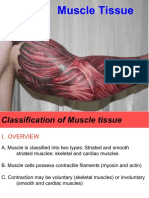
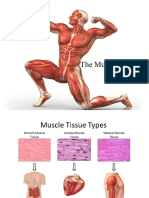
You might also like
- Muscle PhysiologyDocument12 pagesMuscle PhysiologyAyres EvangNo ratings yet
- Chapter 6Document2 pagesChapter 6Nona PeriarceNo ratings yet
- Faal Otot Dan GerakDocument46 pagesFaal Otot Dan GerakElsya Nur FadhillaNo ratings yet
- 589bc774 Df1d 47c6 Bc97 b5fd73030ca6Muscle PhysiologyDocument3 pages589bc774 Df1d 47c6 Bc97 b5fd73030ca6Muscle PhysiologyMalti SharmaNo ratings yet
- Histo MuscleDocument11 pagesHisto MuscleaikaNo ratings yet
- Chapter 9 Actin FloresDocument39 pagesChapter 9 Actin FloresPATRICK MARK FLORESNo ratings yet
- BIOL 2401 Human Anatomy - Phys I - 7.4-7.5 Muscle PhysiologyDocument20 pagesBIOL 2401 Human Anatomy - Phys I - 7.4-7.5 Muscle PhysiologyKevin WhitesideNo ratings yet
- Muscular System: Properties of Muscle TissuesDocument6 pagesMuscular System: Properties of Muscle TissuesaudreyNo ratings yet
- MuscleDocument11 pagesMuscleJeff ParkNo ratings yet
- Muscle ContractionDocument44 pagesMuscle ContractionIshani ManivannanNo ratings yet
- St. Luke's College of Medicine - William H. Quasha Memorial: PhysiologyDocument3 pagesSt. Luke's College of Medicine - William H. Quasha Memorial: PhysiologyMavic VillanuevaNo ratings yet
- Cytoskeleton MADocument22 pagesCytoskeleton MAsema raraNo ratings yet
- Contraction of Skeletal MuscleDocument64 pagesContraction of Skeletal MuscleFhrecy Ann ConsularNo ratings yet
- Excitable Tissues 1Document8 pagesExcitable Tissues 1gojaisaiah787No ratings yet
- Stevens & Lowe's Human Histology 4 EdDocument13 pagesStevens & Lowe's Human Histology 4 EdsheldonboyNo ratings yet
- Chapter 10Document5 pagesChapter 10Nona PeriarceNo ratings yet
- General Mechanism of Muscle ContractionDocument22 pagesGeneral Mechanism of Muscle ContractionAnuya Kulkarni-DeoNo ratings yet
- CH 39 MusclesDocument21 pagesCH 39 MuscleserichaasNo ratings yet
- Skeletal Muscle Contraction and The Motor UnitsDocument4 pagesSkeletal Muscle Contraction and The Motor UnitsAnonymous vnv6QFNo ratings yet
- Muscular System OverviewDocument10 pagesMuscular System OverviewOliver MianaNo ratings yet
- Reviewer - Muscular SystemDocument11 pagesReviewer - Muscular SystemIvy Jan OcateNo ratings yet
- Muscle PhysiologyDocument88 pagesMuscle PhysiologyAftab MughalNo ratings yet
- Chapter 10: Muscle Tissue: VoluntaryDocument13 pagesChapter 10: Muscle Tissue: VoluntaryClyde BaltazarNo ratings yet
- Skeletal Muscle Classification and StructureDocument43 pagesSkeletal Muscle Classification and StructureHusnain WattoNo ratings yet
- Functional Anatomy - ContentDocument14 pagesFunctional Anatomy - ContentxanaduNo ratings yet
- Actin Myosin CytoskeletonDocument28 pagesActin Myosin Cytoskeletonjesus rajeshdivyaNo ratings yet
- Muscle PhysiologyDocument24 pagesMuscle PhysiologyArmando AlehandroNo ratings yet
- Physiology of Skeletal Muscle and Muscle FibreDocument7 pagesPhysiology of Skeletal Muscle and Muscle FibreNoor-E-Khadiza ShamaNo ratings yet
- 46 MusclesDocument5 pages46 Musclesuddim004100% (2)
- Lec Activity7 Muscular System CastroDocument7 pagesLec Activity7 Muscular System CastropelielNo ratings yet
- Muscular TDocument18 pagesMuscular TSumayya KabeerNo ratings yet
- Wa0038Document10 pagesWa0038Shehzad QureshiNo ratings yet
- STRUCTURE OF SKELETAL MUSCLEDocument3 pagesSTRUCTURE OF SKELETAL MUSCLEkrystal1994No ratings yet
- Microscopic Anatomy of A Skeletal Muscle FiberDocument33 pagesMicroscopic Anatomy of A Skeletal Muscle FiberKumar MSNo ratings yet
- 3D. Muscle TissueDocument47 pages3D. Muscle Tissuejaf123xyNo ratings yet
- Célula Muscular, Macro y MicrobiologicoDocument11 pagesCélula Muscular, Macro y MicrobiologicoimanolNo ratings yet
- Muscular System Student Notes With Muscle ListDocument13 pagesMuscular System Student Notes With Muscle Listpham vanNo ratings yet
- UNOANAPHYDocument45 pagesUNOANAPHYGraye SolliNo ratings yet
- Skeletal MusclesDocument7 pagesSkeletal MusclesAhmed OudahNo ratings yet
- Physiology Slide #1Document18 pagesPhysiology Slide #1anon_974423120No ratings yet
- St. Luke's College of Medicine - William H. Quasha Memorial: AnatomyDocument6 pagesSt. Luke's College of Medicine - William H. Quasha Memorial: AnatomyMavic VillanuevaNo ratings yet
- The Muscular SystemDocument9 pagesThe Muscular SystemKathleenJoyGalAlmasinNo ratings yet
- Muscle System 2019Document32 pagesMuscle System 2019ZNA EntertainmentNo ratings yet
- CytoskeletonDocument27 pagesCytoskeletonserenity limNo ratings yet
- B Muscle Tissue HistologyDocument55 pagesB Muscle Tissue HistologyAffie SaikolNo ratings yet
- Muscular SystemDocument6 pagesMuscular SystemTricia Mae LlerenaNo ratings yet
- Wk3 - Muscle Structure, Function and Movement - Student Version 2023Document42 pagesWk3 - Muscle Structure, Function and Movement - Student Version 2023Alexandra GutrovaNo ratings yet
- Muscular System Explained: Skeletal, Cardiac & Smooth Muscle TissueDocument44 pagesMuscular System Explained: Skeletal, Cardiac & Smooth Muscle TissueClaudine Jo B. TalabocNo ratings yet
- Muscular TissuesDocument46 pagesMuscular TissuesDAVE CANALETANo ratings yet
- Muscular Tissue Anatomy and Physiology LectureDocument25 pagesMuscular Tissue Anatomy and Physiology LecturehanzNo ratings yet
- Tortora Outline Chapter 10 For PPDocument12 pagesTortora Outline Chapter 10 For PPTasha BradyNo ratings yet
- Skeletal Muscle LecturesDocument14 pagesSkeletal Muscle Lecturesanis rahmawatiNo ratings yet
- Muscular SystemDocument9 pagesMuscular SystemReynaldo MontañoNo ratings yet
- Skeletal MuscleDocument2 pagesSkeletal Musclesteev spideyNo ratings yet
- Muscle Tissue LectureDocument34 pagesMuscle Tissue LectureurjaNo ratings yet
- Module5 Muscular SystemDocument28 pagesModule5 Muscular Systemwilden ellorincoNo ratings yet
- Physiology of Skeletal & Smooth MusclesDocument10 pagesPhysiology of Skeletal & Smooth MusclesAli AlhamadNo ratings yet
- 7 Muscular SystemDocument11 pages7 Muscular SystemmargaretNo ratings yet
- Classification of MusclesDocument50 pagesClassification of MusclesNoor HaleemNo ratings yet
- Case Digest - Reyes Vs Sisters of Mercy HospitalDocument2 pagesCase Digest - Reyes Vs Sisters of Mercy HospitalLorenz L. Llamas IIINo ratings yet
- Case Digest - Carillo Vs PeopleDocument2 pagesCase Digest - Carillo Vs PeopleLorenz L. Llamas IIINo ratings yet
- Carillo V PeopleDocument10 pagesCarillo V PeopleLorenz L. Llamas IIINo ratings yet
- Solidum Et Al vs. People G.R. No. 192123 March 10, 2014Document18 pagesSolidum Et Al vs. People G.R. No. 192123 March 10, 2014FD BalitaNo ratings yet
- Garcia-Rueda v. Office of The City ProsecutorDocument5 pagesGarcia-Rueda v. Office of The City ProsecutorLorenz L. Llamas IIINo ratings yet
- 03c Conditions Leading To Muscle WeaknessDocument5 pages03c Conditions Leading To Muscle WeaknessLorenz L. Llamas IIINo ratings yet
- G.R. No. 166097Document8 pagesG.R. No. 166097Lorenz L. Llamas IIINo ratings yet
- G.R. No. 178763Document16 pagesG.R. No. 178763Lorenz L. Llamas IIINo ratings yet
- Chapter VI - Contraction of The Skeletal Muscle: Filamentous Bands Light Bands Dark BandsDocument13 pagesChapter VI - Contraction of The Skeletal Muscle: Filamentous Bands Light Bands Dark BandsLorenz L. Llamas IIINo ratings yet
- GoutDocument3 pagesGoutLorenz L. Llamas IIINo ratings yet
- Human Physiology Lecture 1Document10 pagesHuman Physiology Lecture 1Lorenz L. Llamas IIINo ratings yet
- Biology Master PDFDocument7 pagesBiology Master PDFRae Krystylynne CodillaNo ratings yet
- Philippine National Formulary 8thED PDFDocument800 pagesPhilippine National Formulary 8thED PDFGio Mari Marcial100% (3)
- Human Physiology Lecture 1Document10 pagesHuman Physiology Lecture 1Lorenz L. Llamas IIINo ratings yet
- PNF Eml1Document57 pagesPNF Eml1SheinGervacioErnacioNo ratings yet
- Cip On CovidDocument40 pagesCip On CovidLorenz L. Llamas IIINo ratings yet
- Pharmacovigilance 13 Nov 2019 DOHDocument28 pagesPharmacovigilance 13 Nov 2019 DOHLorenz L. Llamas IIINo ratings yet
- Biology Master PDFDocument7 pagesBiology Master PDFRae Krystylynne CodillaNo ratings yet
- Philippine Antibiotic Awareness Week 2019Document27 pagesPhilippine Antibiotic Awareness Week 2019Lorenz L. Llamas IIINo ratings yet
- 2 DOH Program To Combat AMRDocument15 pages2 DOH Program To Combat AMRLorenz L. Llamas IIINo ratings yet
- Biology Master PDFDocument7 pagesBiology Master PDFRae Krystylynne CodillaNo ratings yet
- 3rd Quarter Biology Note Grade 10Document7 pages3rd Quarter Biology Note Grade 10G and B Family buildingNo ratings yet
- Evolution EvidenceDocument7 pagesEvolution EvidenceMiljoy DelegadoNo ratings yet
- Study of The Influence of Electromagnetic Fields On Human Well-Being in Buildings of The Urban Area of Annaba (Algeria)Document9 pagesStudy of The Influence of Electromagnetic Fields On Human Well-Being in Buildings of The Urban Area of Annaba (Algeria)International Network For Natural SciencesNo ratings yet
- 2 Pitutary GlandDocument44 pages2 Pitutary GlandHanen ZedanNo ratings yet
- SEP 2023 Syllabus Human Physiology Updated 5-29-23Document6 pagesSEP 2023 Syllabus Human Physiology Updated 5-29-23gfsgfdsgfdNo ratings yet
- Insights Into Insecticide-Resistance Mechanisms in Invasive SpeciesDocument18 pagesInsights Into Insecticide-Resistance Mechanisms in Invasive SpeciesNg Kin HoongNo ratings yet
- DR - REVATI - IMMUNE MEDIATED DISEASES OF OROFACIAL REGION - RevatiDocument72 pagesDR - REVATI - IMMUNE MEDIATED DISEASES OF OROFACIAL REGION - RevatiNanik AndianiNo ratings yet
- ESSENTIALS OF HUMAN EMBRYOLOGY by Prof NGASSAPADocument315 pagesESSENTIALS OF HUMAN EMBRYOLOGY by Prof NGASSAPAJumbe Mohamed100% (3)
- Module 4Document37 pagesModule 4Jeraline PiezasNo ratings yet
- Head To Toe Checklist OlderDocument1 pageHead To Toe Checklist Olderspatrick32100% (4)
- Incorporation of The Natural Marine Multi Mineral Dietary Supplement Aquamin Enhances Osteogenesis and Improves The MechanicalDocument10 pagesIncorporation of The Natural Marine Multi Mineral Dietary Supplement Aquamin Enhances Osteogenesis and Improves The Mechanicalana mariaNo ratings yet
- Photosynthesis and RespirationDocument46 pagesPhotosynthesis and RespirationCarmsNo ratings yet
- The Corrs - UnpluggedDocument4 pagesThe Corrs - UnpluggedAnn ArnstbergNo ratings yet
- Chapter 35Document9 pagesChapter 35Jyothis James100% (2)
- Therapeutic Exercise: Foundation & Techniques: Resource Person: Dr. Rahat Ayub PT SHS.326.Lec.13Document30 pagesTherapeutic Exercise: Foundation & Techniques: Resource Person: Dr. Rahat Ayub PT SHS.326.Lec.13Mustafa ,GhulamNo ratings yet
- NEUROMUSCULAR TRANSMISSION: ANATOMY AND PHYSIOLOGYDocument45 pagesNEUROMUSCULAR TRANSMISSION: ANATOMY AND PHYSIOLOGYparuNo ratings yet
- Introduction To Gerontology and Theories of AgingDocument106 pagesIntroduction To Gerontology and Theories of AgingCyden Shame delos Santos100% (1)
- This Content Downloaded From 184.176.223.154 On Mon, 17 Apr 2023 16:11:12 UTCDocument13 pagesThis Content Downloaded From 184.176.223.154 On Mon, 17 Apr 2023 16:11:12 UTCJACKSON LOWNo ratings yet
- The Integumentary SystemDocument4 pagesThe Integumentary SystemKhan Hayudini SaliNo ratings yet
- Dr. Subhasis Roy: Consultant, Sisu Sanjiban Hospital, Salt Lake, KolkataDocument19 pagesDr. Subhasis Roy: Consultant, Sisu Sanjiban Hospital, Salt Lake, Kolkatacy_alcantaraNo ratings yet
- Pathology For The Physical Therapist Assistant 1st Edition Lescher Test BankDocument14 pagesPathology For The Physical Therapist Assistant 1st Edition Lescher Test Bankturnmiddayu0ppsx100% (27)
- Unit 9 - Pain ManagementDocument36 pagesUnit 9 - Pain ManagementPetrinaNo ratings yet
- Calcium Ion As Cellular Messenger: Science ChinaDocument5 pagesCalcium Ion As Cellular Messenger: Science ChinaAhmad Ligar SuhermanNo ratings yet
- Act 2 PDFDocument6 pagesAct 2 PDFKhara TeanoTanNo ratings yet
- 15 - Supportive Care and Emergency TherapyDocument35 pages15 - Supportive Care and Emergency TherapyPeterNo ratings yet
- How Anthropic Noise Impacts Insect Behavior and InteractionsDocument12 pagesHow Anthropic Noise Impacts Insect Behavior and InteractionsJuan LópezNo ratings yet
- Neuro Exam Skill SheetDocument2 pagesNeuro Exam Skill SheetMuhammed ElgasimNo ratings yet
- HENGG-5321, Ethylene & RespirationDocument12 pagesHENGG-5321, Ethylene & RespirationbhagwatNo ratings yet
- 1 s2.0 S0828282X22001271 MainDocument11 pages1 s2.0 S0828282X22001271 MainLilianne Mbengani LaranjeiraNo ratings yet
- Chapter 14 - Multiple ChoiceDocument13 pagesChapter 14 - Multiple ChoiceNicky PhakathiNo ratings yet